The Critical Role of Nurr1 as a Mediator and Therapeutic Target in Alzheimer’s Disease-related Pathogenesis
Jeon Seong Gak1, Yoo Anji1, Chun Dong Wook1, Hong Sang Bum1, Chung Hyunju2, Kim Jin-il3, *, Moon Minho1, *
1Department of Biochemistry, College of Medicine, Konyang University, Daejeon, 35365, Republic of Korea2Department of Core Research Laboratory, Clinical Research Institute, Kyung Hee University Hospital at Gangdong, Seoul 05278, Republic of Korea3Department of Nursing, College of Nursing, Jeju National University, Jeju-si 63243, Republic of Korea
通讯作者: Correspondence should be addressed to: Dr. Minho Moon, College of Medicine, Konyang University, Republic of Korea. Email: hominmoon@konyang.ac.kr and Jin-il Kim, College of Nursing, Jeju National University, Republic of Korea. Email: neoreva@hanmail.net. #These authors contributed equally to this work.Correspondence should be addressed to: Dr. Minho Moon, College of Medicine, Konyang University, Republic of Korea. Email: hominmoon@konyang.ac.kr and Jin-il Kim, College of Nursing, Jeju National University, Republic of Korea. Email: neoreva@hanmail.net. #These authors contributed equally to this work.Correspondence should be addressed to: Dr. Minho Moon, College of Medicine, Konyang University, Republic of Korea. Email: hominmoon@konyang.ac.kr and Jin-il Kim, College of Nursing, Jeju National University, Republic of Korea. Email: neoreva@hanmail.net. #These authors contributed equally to this work.
收稿日期:2019-05-14
修回日期: 2019-06-18
接受日期: 2019-07-18
网络出版日期: 2020-05-09
版权声明:
2020 This is an Open Access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
展开
Abstract
Several studies have revealed that the transcription factor nuclear receptor related 1 (Nurr1) plays several roles not only in the regulation of gene expression related to dopamine synthesis, but also in alternative splicing, and miRNA targeting. Moreover, it regulates cognitive functions and protects against inflammation-induced neuronal death. In particular, the role of Nurr1 in the pathogenesis of Parkinson’s disease (PD) has been well investigated; for example, it has been shown that it restores behavioral and histological impairments in PD models. Although many studies have evaluated the connection between Nurr1 and PD pathogenesis, the role of Nurr1 in Alzheimer’s disease (AD) remain to be studied. There have been several studies describing Nurr1 protein expression in the AD brain. However, only a few studies have examined the role of Nurr1 in the context of AD. Therefore, in this review, we highlight the overall effects of Nurr1 under the neuropathologic conditions related to AD. Furthermore, we suggest the possibility of using Nurr1 as a therapeutic target for AD or other neurodegenerative disorders.
JeonSeong Gak, YooAnji, ChunDong Wook, HongSang Bum, ChungHyunju, KimJin-il, MoonMinho. The Critical Role of Nurr1 as a Mediator and Therapeutic Target in Alzheimer’s Disease-related Pathogenesis[J]. Aging and Disease, 2020, 11(3): 705-724 https://doi.org/10.14336/AD.2019.0718
1. The nuclear receptor related-1 protein, Nurr1
The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12].
In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20].
Table 1
Table 1 Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1).
2.1. Neuroprotective effects of Nurr1
Neuronal death is the main pathogenic factor underlying neurodegenerative diseases such as AD and PD [67, 68]. Under neuropathological conditions, including neuroinflammation, excitotoxicity, and oxidative stress, neurons can be rescued by upregulation or activation of neuroprotective factors such as reactive oxygen species (ROS) scavengers or anti-apoptotic molecules [69-71]. Therefore, induction of neuroprotective factors may be a therapeutic strategy for the treatment of neuro-degeneration-related diseases.
Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16].
2.2. Anti-inflammatory effects of Nurr1
Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells.
2.3. Peripheral immune cell modulation of Nurr1
The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106].
Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115].
Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD.
2.4. Cell-cycle regulation of Nurr1 and AD
Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133].
The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18].
These results suggest that Nurr1 may interfere negatively or positively with the cell cycle depending on the cell type and its environment. They also suggest that Nurr1 may have a positive effect on AD by promoting proliferation of neural stem cells or by suppressing the abnormally promoted cell cycle in the AD brain. However, the correlation of Nurr1 with the cell cycle in AD has not yet been directly reported.
2.5. Neurogenic effects of Nurr1 in the adult brain
Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148].
A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16].
2.6. Memory-enhancing effects of Nurr1
Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions.
Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170].
Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD.
2.7. The role of Nurr1 in vascular pathologies
Cerebral amyloid angiopathy (CAA) is an AD-related histopathology showing pathological changes such as deposits of Aβ in the blood vessels of the CNS and the walls of leptomeningeal arteries [173]. CAA is not only associated with the Aβ burden of the brain parenchyma, but also occasionally induces necrosis resulting in cerebral hemorrhage [174, 175]. However, the degree of CAA varies between AD brains, and the majority of AD patients showing microvascular amyloid deposits do not experience cerebral hemorrhage [176]. Nonetheless, because CAA could contribute to cortical dysfunction, regulation of vascular Aβ accumulation is suggested for mitigation of secondary AD pathology.
In human atherosclerosis, expression of NR4A including Nurr1 receptors is increased in macrophages after inflammatory stimulation [177]. In addition, overexpression of Nurr1 inhibited vascular lesion formation through reduction of smooth muscle cell proliferation and inflammatory response [135, 178]. These findings prove that endogenous Nurr1 suppresses macrophage activation, foam-cell formation, and further differentiation. Thus, it provides further evidence that atherogenesis could be prevented by modulation of Nurr1 expression. Therefore, Nurr1 may be proposed as a novel therapeutic target for preventing vascular disruption-related diseases mediated by AD.
2.8. The role of Nurr1 in metabolism
There are a number of studies describing both a correlation between type 2 diabetes and the risk of AD, and an association of hyperglycemia with AD pathophysiology [179, 180]. In addition, it has been reported that excess weight in middle age is related to increased risk of AD [181]. Metabolic hormones such as insulin, leptin, ghrelin, and adiponectin have been reported to have therapeutic potential and are also involved in the pathogenesis of AD [182]. Since AD could be accompanied by metabolic disturbances, normalization of metabolism will provide new insights into AD treatment.
There are several studies showing the roles of Nr4a receptors associated with metabolic functions. In particular, it has been reported that aberrant expression of Nurr1 is correlated to glucose metabolism and metabolic syndrome [183, 184]. Moreover, Nr4a receptors are expressed under metabolism-related stimuli including cold, fatty acids, glucose, insulin, and cholesterol. Therefore, it has been suggested that Nr4a receptors can be therapeutic targets for metabolism-related disorders [185]. There is also compelling evidence suggesting that Nr4a receptors show a potent association with the course of type 2 diabetes through regulation of insulin sensitivity and glucose homeostasis. Even though the mechanisms underlying the regulation of glucose metabolism by Nr4a receptors have not been elucidated, Nr4a receptors are identified as potential biological targets for diabetic patients [186]. However, the association of Nurr1 with carbohydrate metabolism is currently unclear. Therefore, identification of the correlation between Nurr1 and metabolic disorders in AD will be a novel pioneering field of study.
2.8. The role of Nurr1 in the habenula
The habenula is a part of the epithalamus in the diencephalon, located dorsal-medial to the posterior thalamus. It is divided into two regions: (1) the lateral habenula (LHb), which is innervated by the rostromedial tegmental nucleus (RMTg) to the dopamine-related regions, such as the ventral tegmental area (VTA) and substantia nigra (SN) or the serotonin system, such as the dorsal raphe nucleus and median raphe nucleus, and (2) the medial habenula (MHb), which is innervated with the serotonin system through the interpeduncular nucleus (IPN) [187-190]. The habenula, which is connected to the limbic system, basal ganglia, and pineal gland, is involved in the reward system and cognitive functions such as learning, memory, and attention [191]. Therefore, it has been suggested that the habenula may be involved in psychiatric disorders such as depression, schizophrenia, and drug-induced psychosis [192].
In adults and during development, the Nurr1 and Nr4a2 genes are robustly and specifically expressed in the MHb expressing both the choline acetyltransferase in the ventral part and the neuropeptide SP in the dorsal part [191, 193-195]. In addition, Nurr1 has been reported to mediate a gene pathway involved in habenula development regulated by the POU-domain transcription factor Brn3a [195]. Notably, several studies have suggested that habenular activity is involved in depression, which is one of the most common psychiatric symptoms in AD and a risk factor for AD development.
In the genetic helpless model and α-methyl-para-tyrosine-induced depression model, brain metabolism and glucose metabolism were elevated in the habenular compared to the control while metabolism of other brain regions was reduced [196, 197]. In patients with depression, it is shown that habenula activity is strongly correlated with dorsal raphé nuclei activity providing an evidence for the important roles of feedback pathway between habenula and dorsal raphé nuclei in controlling release of serotonin [198]. Application of deep brain stimulation to the LHb in a therapy-refractory patient with depression caused successful remission of depression, clarifying the relationship between the habenula and depression [199]. These reports suggest that upregulation of Nurr1 may have a positive effect on not only AD-related pathology but also the psychiatric symptoms that may occur in patients with AD.
3. Therapeutic potential of Nurr1 activation for AD treatment
As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16].
Interestingly, Nurr1 not only forms homo- or heterodimers with other members of the NR4A family, but can also forms heterodimers with RXRs via DR5 response element [201]. In addition, since the activation of nuclear receptors such as RXR, liver X receptor (LXR), and peroxisome proliferator-activated receptor (PPAR)-γ is known to affect Aβ generation and Aβ clearance, Nurr1 may contribute to the alleviation of Aβ-related pathophysiology through interactions with other nuclear receptors [202-205].
Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16].
Figure 1. Overview of effect of Nurr1 in Alzheimer’s disease.
4. Nurr1 and brain disorders
As mentioned earlier, Nurr1 can contribute to the beneficial effects on AD-associated pathologies and may have therapeutic potential for AD. Notably, there have been studies regarding the roles of Nurr1 in various conditions in related to AD pathologies. It is speculated that reviewing the roles of Nurr1 in these conditions could provide and extend insights about possible applications of Nurr1 in the modulation of AD pathogenesis. In the following sections, the roles of Nurr1 in brain disorders will be described.
4.1. Parkinson’s disease
Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy.
4.2. Ischemic stroke
A recent study has shown that Nurr1 expression was dynamic following acute ischemia induced by middle cerebral artery occlusion (MCAO)/reperfusion in a rat model. There was a negative correlation between Nurr1 and infarct volume up to 12 hours after ligation, but a positive correlation was observed after 24 hours. Nurr1 overexpression inhibited tumor necrosis factor-α (TNF-α) levels in microglia. Increase of Nurr1 expression through suppression of miR-145-5p, a negative regulator of Nurr1 alleviated infarct volume and improved the neurological outcomes in an acute stroke model [223]. In addition, transplantation of Nurr1-overexpressing human embryonal carcinoma cells into the ischemic striatum restored the behavioral disorder in a transient MCAO rat model [153]. Considering that TNF-α released from Aβ-activated microglia is a key cytokine causing cell cycle events, which are related to pathogenesis of neuronal death in AD [224], these regulatory effects of Nurr1 on microglial activation could be a therapeutic target for AD as well as ischemic stroke.
4.3. Schizophrenia
Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases.
4.4. Addictive behaviors
Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors.
VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis.
4.6. Circadian rhythm disorder
Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD.
5. Conclusion
Recent findings regarding the Nurr1 role in the CNS have demonstrated molecular, cellular, and physiological responses underlying various conditions, and these findings may provide insights for the association between Nurr1 and the underlying mechanisms of AD (Fig. 1). Studies on the effect of Nurr1 support the correlation between Nurr1 expression and various stages of AD pathology and symptoms, including neuronal cell death, inflammation, synaptic loss, impaired adult neurogenesis, psychiatric symptoms, and cognitive deficits.
All mechanisms of development of neuro-degenerative diseases, especially of AD, are closely related to the actions of Nurr1. Nurr1 may be capable of regulating AD-related pathogenesis, based on recent studies showing the critical roles of Nurr1 in AD-related pathology (Fig. 1). As a result, subsequent experiments have been performed to prove Nurr1 as a potential target for treatment of AD, and have suggested Nurr1 agonists/mimetics as potential therapeutic agents for AD.
Conflict of interest
The authors have no conflict of interest to declare.
Acknowledgments
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2018R1D1A3B07041059 to M.M and 2017R1D1A1B03035839 to H.C) and by the Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01319901 and PJ01428603), Rural Development Administration, Republic of Korea
Cellular expression of the immediate early transcription factors Nurr1 and NGFI-B suggests a gene regulatory role in several brain regions including the nigrostriatal dopamine system
SmithGA, RochaEM, RooneyT, BarneoudP, McLeanJR, BeaganJ, et al. (2015).
A Nurr1 agonist causes neuroprotection in a Parkinson's disease lesion model primed with the toll-like receptor 3 dsRNA inflammatory stimulant poly(I:C)
BarkerWW, LuisCA, KashubaA, LuisM, HarwoodDG, LoewensteinD, et al. (2002).
Relative frequencies of Alzheimer disease, Lewy body, vascular and frontotemporal dementia, and hippocampal sclerosis in the State of Florida Brain Bank
MoonM, HongHS, NamDW, BaikSH, SongH, KookSY, et al. (2012).
Intracellular amyloid-beta accumulation in calcium-binding protein-deficient neurons leads to amyloid-beta plaque formation in animal model of Alzheimer's disease
LinX, ParisiadouL, SgobioC, LiuG, YuJ, SunL, et al. (2012).
Conditional expression of Parkinson's disease-related mutant alpha-synuclein in the midbrain dopaminergic neurons causes progressive neurodegeneration and degradation of transcription factor nuclear receptor related 1
Hypoxia enhances stimulating effect of amyloid beta peptide (25-35) for interleukin 17 and T helper lymphocyte subtype 17 upregulation in cultured peripheral blood mononuclear cells
IFN-gamma Production by amyloid beta-specific Th1 cells promotes microglial activation and increases plaque burden in a mouse model of Alzheimer's disease
. J Immunol, 190:2241-2251.
[104]
ZhangJ, KeKF, LiuZ, QiuYH, PengYP (2013).
Th17 cell-mediated neuroinflammation is involved in neurodegeneration of abeta1-42-induced Alzheimer's disease model rats
Lambracht-WashingtonD, QuBX, FuM, AndersonLD, Jr., StuveO, EagarTN, et al. (2011).
DNA immunization against amyloid beta 42 has high potential as safe therapy for Alzheimer's disease as it diminishes antigen-specific Th1 and Th17 cell proliferation
Towards understanding the role of orphan nuclear receptor NR4A2 in Th17 cell-mediated central nervous system autoimmunity: An experimental approach using an animal model of multiple sclerosis
. Clinical and Experimental Neuroimmunology, 5:137-148.
Modulation of orphan nuclear receptor NURR1 expression by methotrexate in human inflammatory joint disease involves adenosine A2A receptor-mediated responses
BontaPI, PolsTW, van TielCM, VosM, ArkenboutEK, RohlenaJ, et al. (2010).
Nuclear receptor Nurr1 is expressed in and is associated with human restenosis and inhibits vascular lesion formation in mice involving inhibition of smooth muscle cell proliferation and inflammation
Vergano-VeraE, Diaz-GuerraE, Rodriguez-TraverE, Mendez-GomezHR, SolisO, PignatelliJ, et al. (2015).
Nurr1 blocks the mitogenic effect of FGF-2 and EGF, inducing olfactory bulb neural stem cells to adopt dopaminergic and dopaminergic-GABAergic neuronal phenotypes
ShimJW, ParkCH, BaeYC, BaeJY, ChungS, ChangMY, et al. (2007).
Generation of functional dopamine neurons from neural precursor cells isolated from the subventricular zone and white matter of the adult rat brain using Nurr1 overexpression
HaraK, MatsukawaN, YasuharaT, XuL, YuG, MakiM, et al. (2007).
Transplantation of post-mitotic human neuroteratocarcinoma-overexpressing Nurr1 cells provides therapeutic benefits in experimental stroke: in vitro evidence of expedited neuronal differentiation and GDNF secretion
Proneural bHLH neurogenin 2 differentially regulates Nurr1-induced dopamine neuron differentiation in rat and mouse neural precursor cells in vitro
. FEBS Lett, 582:537-542.
[155]
ChenX, QianY, WangX, TangZ, XuJ, LinH, et al. (2018).
Nurr1 promotes neurogenesis of dopaminergic neuron and represses inflammatory factors in the transwell coculture system of neural stem cells and microglia
ClarkeSE, JukesMC, NjagiJK, KhasakhalaL, CundillB, OtidoJ, et al. (2008).
Effect of intermittent preventive treatment of malaria on health and education in schoolchildren: a cluster-randomised, double-blind, placebo-controlled trial
The mesopontine rostromedial tegmental nucleus: A structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta
Mechanisms underlying the rapid peroxisome proliferator-activated receptor-gamma-mediated amyloid clearance and reversal of cognitive deficits in a murine model of Alzheimer's disease
. J Neurosci, 32:10117-10128.
[205]
Mandrekar-ColucciS, LandrethGE (2011).
Nuclear receptors as therapeutic targets for Alzheimer's disease
CastilloSO, BaffiJS, PalkovitsM, GoldsteinDS, KopinIJ, WittaJ, et al. (1998).
Dopamine biosynthesis is selectively abolished in substantia nigra/ventral tegmental area but not in hypothalamic neurons in mice with targeted disruption of the Nurr1 gene
Heroin abuse is characterized by discrete mesolimbic dopamine and opioid abnormalities and exaggerated nuclear receptor-related 1 transcriptional decline with age
Rhynchophylline Downregulates Phosphorylated cAMP Response Element Binding Protein, Nuclear Receptor-related-1, and Brain-derived Neurotrophic Factor Expression in the Hippocampus of Ketamine-induced Conditioned Place Preference Rats
LoweN, KirleyA, HawiZ, ShamP, WickhamH, KratochvilCJ, et al. (2004).
Joint analysis of the DRD5 marker concludes association with attention-deficit/hyperactivity disorder confined to the predominantly inattentive and combined subtypes
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
... ]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Structure and function of Nurr1 identifies a class of ligand-independent nuclear receptors
1
2003
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Cellular expression of the immediate early transcription factors Nurr1 and NGFI-B suggests a gene regulatory role in several brain regions including the nigrostriatal dopamine system
1
1996
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Comparative distribution of NURR1 and NUR77 nuclear receptors in the mouse central nervous system
1
1996
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
The function and mechanisms of Nurr1 action in midbrain dopaminergic neurons, from development and maintenance to survival
1
2012
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
The role of Nurr1 in the development of dopaminergic neurons and Parkinson's disease
1
2005
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons
2
1998
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Multiple signaling pathways regulate the transcriptional activity of the orphan nuclear receptor NURR1
1
2006
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Long 3'UTR of Nurr1 mRNAs is targeted by miRNAs in mesencephalic dopamine neurons
1
2017
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Bipartite functions of the CREB co-activators selectively direct alternative splicing or transcriptional activation
1
2009
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
CRTC1 mediates preferential transcription at neuronal activity-regulated CRE/TATA promoters
1
2017
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
The NR4A subgroup: Immediate early response genes with pleiotropic physiological roles
1
2006
... The nuclear receptor related 1 (Nurr1) protein, also known as nuclear receptor subfamily 4, group A, member 2 (NR4A2). Nurr1 belongs to the nuclear receptor subfamily 4A (NR4A), which consists of NR4A1, NR4A2, and NR4A3, also known as Nur77, Nurr1, and Nor1, respectively [1, 2]. Nurr1 is robustly expressed in the central nervous system (CNS) [3, 4]. Similar to other members of the NR4A, Nurr1 has been considered as an orphan nuclear receptor, whose endogenous ligand has not been identified [1]. Nurr1 is well known to play an essential role in the development, function, and maintenance of midbrain dopaminergic neurons [5-7]. In particular, Nurr1 is known to play an integral role in multiple signaling pathways involved in the differentiation and phenotype of dopaminergic neurons [8]. It is also targeted by miRNAs in dopaminergic neurons, and is alternatively spliced by cyclic adenosine monophosphate (cAMP)-responsive element-binding protein (CREB)-regulated transcription co-activators [9, 10]. Notably, a recent report suggesting that CREB-regulated transcription coactivator-1 (CRTC1) mediates expression of the Nurr1 gene provided evidence for specific molecular mechanisms for the regulation of Nurr1 expression in primary cortical neurons [11]. Nurr1 expression is also found to be directly induced by various stimuli such as inflammatory signals. Once Nurr1 is activated by various factors, it binds to specific DNA sequences in the promoter region of the target genes to positively regulate their expression [12]. ...
Nurr1 in Parkinson's disease and related disorders
6
2006
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
... ]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
... ]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Decreased ethanol preference and wheel running in Nurr1-deficient mice
2
2003
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Adult mice with reduced Nurr1 expression: an animal model for schizophrenia
7
2007
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
... Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
... ] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
Nurr1 (NR4A2) regulates Alzheimer's disease-related pathogenesis and cognitive function in the 5XFAD mouse model
11
2018
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
... ], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
The Nurr1 Ligand,1,1-bis(3'-Indolyl)-1-(p-Chlorophenyl)Methane, Modulates Glial Reactivity and Is Neuroprotective in MPTP-Induced Parkinsonism
4
2018
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... ]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
The pharmacological stimulation of Nurr1 improves cognitive functions via enhancement of adult hippocampal neurogenesis
5
2016
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
Nuclear receptor Nurr1 agonists enhance its dual functions and improve behavioral deficits in an animal model of Parkinson's disease
6
2015
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... , 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
A Nurr1 agonist causes neuroprotection in a Parkinson's disease lesion model primed with the toll-like receptor 3 dsRNA inflammatory stimulant poly(I:C)
6
2015
... In addition to the role of Nurr1 in the pathogenesis of dopamine-related neurological disorders [6, 13], several studies have revealed the involvement of Nurr1 in reward-seeking behavior [14], symptoms of schizophrenia [15], and pathogenesis of Alzheimer’s disease (AD) [16]. Although the endogenous ligand of Nurr1 has not yet been identified, the cognition-enhancing effects of Nurr1 agonists, which have been demonstrated in wild-type (WT) and PD mice, support the potential of Nurr1 as a therapeutic target for neurodegenerative disease [17-20]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... , 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Relative frequencies of Alzheimer disease, Lewy body, vascular and frontotemporal dementia, and hippocampal sclerosis in the State of Florida Brain Bank
1
2002
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer's disease: the challenge of the second century
1
2011
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer's disease and senile dementia: loss of neurons in the basal forebrain
1
1982
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Heterogeneity and selectivity of the degeneration of cholinergic neurons in the basal forebrain of patients with Alzheimer's disease
0
1993
The cholinergic hypothesis of Alzheimer's disease: a review of progress
1
1999
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Smoking and increased Alzheimer's disease risk: a review of potential mechanisms
1
2014
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
A review on Alzheimer's disease pathophysiology and its management: an update
0
2015
Risk factors for Alzheimer's disease: a prospective analysis from the Canadian Study of Health and Aging
0
2002
Pathogenesis of Alzheimer’s disease
1
2007
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer's disease: the size of the problem, clinical manifestation and heterogeneity
1
1998
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Comparison of Clinical Manifestation in Familial Alzheimer's disease and Dementia with Lewy Bodies
1
2008
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Advances in the early detection of Alzheimer's disease
1
2004
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer's Disease
1
2004
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
The genetic landscape of Alzheimer disease: clinical implications and perspectives
1
2016
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Genetic aspects of Alzheimer disease
1
2008
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Acetylcholinesterase inhibitors in Alzheimer’s disease
1
1999
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Cholinesterase inhibitors for Alzheimer's disease
0
2006
The glutamatergic system and Alzheimer's disease: therapeutic implications
0
2003
Memantine and Cholinesterase Inhibitors: Complementary Mechanisms in the Treatment of Alzheimer’s Disease
1
2013
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Solanezumab for the treatment of mild-to-moderate Alzheimer’s disease
1
2012
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
An effector-reduced anti-beta-amyloid (Abeta) antibody with unique abeta binding properties promotes neuroprotection and glial engulfment of Abeta
0
2012
Alzheimer disease: Plaque-specific anti-A[beta] antibody shows promise in model of AD
0
2013
Immunotherapy for Alzheimer's disease: from anti-beta-amyloid to tau-based immunization strategies
1
2012
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer mechanisms and therapeutic strategies
1
2012
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Alzheimer's disease: a central role for amyloid
1
1994
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Intracellular amyloid-beta in Alzheimer's disease
0
2007
Intracellular amyloid-beta accumulation in calcium-binding protein-deficient neurons leads to amyloid-beta plaque formation in animal model of Alzheimer's disease
1
2012
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
The role of NR4A transcription factors in memory formation
4
2011
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Knockdown of Nurr1 in the rat hippocampus: implications to spatial discrimination learning and memory
2
2006
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Hippocampal expression of the orphan nuclear receptor gene hzf-3/nurr1 during spatial discrimination learning
2
2000
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Schizophrenia-relevant behaviors in a genetic mouse model of constitutive Nurr1 deficiency
1
2011
... Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
HDAC3 is a critical negative regulator of long-term memory formation
0
2011
Differential roles for Nr4a1 and Nr4a2 in object location vs. object recognition long-term memory
3
2012
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
NR4A nuclear receptors support memory enhancement by histone deacetylase inhibitors
4
2012
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
The NR4A orphan nuclear receptors mediate transcription-dependent hippocampal synaptic plasticity
4
2013
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
Nurr1 protein is required for N-methyl-D-aspartic acid (NMDA) receptor-mediated neuronal survival
2
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
NR4A orphan nuclear receptors as mediators of CREB-dependent neuroprotection
0
NGFI-B nuclear orphan receptor Nurr1 interacts with p53 and suppresses its transcriptional activity
0
2009
Reduced Nurr1 expression increases the vulnerability of mesencephalic dopamine neurons to MPTP-induced injury
3
1999
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
Microarray analyses support a role for Nurr1 in resistance to oxidative stress and neuronal differentiation in neural stem cells
5
2007
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... ]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... , 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
Toward neuroprotective treatments of Parkinson's disease
1
2017
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
beta-Amyloid disrupts activity-dependent gene transcription required for memory through the CREB coactivator CRTC1
2
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Crtc1 activates a transcriptional program deregulated at early Alzheimer's disease-related stages
1
2014
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
Correlation between orphan nuclear receptor Nurr1 expression and amyloid deposition in 5XFAD mice, an animal model of Alzheimer's disease
2
2015
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
Cellular Model of Alzheimer's Disease: Abeta1-42 Peptide Induces Amyloid Deposition and a Decrease in Topo Isomerase IIbeta and Nurr1 Expression
3
2017
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
... model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Whole transcriptome profiling of Late-Onset Alzheimer's Disease patients provides insights into the molecular changes involved in the disease
1
2018
... AD is known to be the most common cause of dementia and is responsible for 60%-70% of the cases of dementia [21, 22]. AD patients exhibit impairment of cognitive functions, which is mediated by abnormal accumulation of amyloid plaques containing amyloid beta (Aβ) and neurofibrillary tangles (NFT) in the brain [23-25]. With the increasing focus on AD over the past century because of the gradual aging of the global population, the pathophysiologies of AD [26-29], as well as its clinical manifestations [30, 31], diagnosis [32, 33], and genetic characteristics [34, 35] are now relatively well understood. Several studies on therapeutic approaches for AD have been performed, including those involving cholinesterase inhibitors, N-methyl-d-aspartate (NMDA) receptor antagonists [36-39], and anti-Aβ therapy [40-43]. Nevertheless, there is no disease-modifying therapy yet [44]. Although the exact mechanisms of AD pathogenesis are unclear, intracellular and extracellular Aβ are thought to be major causative factors associated with AD-related pathologies, such as neurodegeneration and cognitive dysfunction [45-47]. Interestingly, Nurr1 is known to act as a critical regulator of hippocampal function, hippocampal synaptic plasticity, and cognitive functions [15, 48-55], and is an essential mediator of neuroprotection or anti-inflammation after exposure to neuropathological stress [19, 56-61]. In addition, a number of studies have indicated altered levels of Nurr1 in Aβ-treated neuronal cells, animal models of AD, and the brains of patients with AD [13, 62-65], implying that Nurr1 may play a role in the pathogenesis of AD. Recent studies have shown that Aβ1-42 fibrils not only lead to upregulation of tau hyperphosphorylation and presenilin 1 mRNA, which are hallmarks of AD pathology, but also significantly reduce Nurr1 mRNA levels in an in vitro model of AD [65]. Immunofluorescence staining with Nurr1-specific antibody in 5XFAD mice, an animal model of AD, showed that the Nurr1 protein is markedly expressed in the brain areas with Aβ accumulation. Moreover, the number of Nurr1-expressing cells is decreased in 5XFAD mice with AD progression, compared with WT mice [64]. In contrast, the levels of miR-184, which directly targets the 3′ UTR of the NR4A2 transcript, are reduced in the hippocampus of late-onset AD patients. In addition, expression of NR4A2 and miR-184 is inversely correlated [66]. These findings suggest that Nurr1 is not only highly implicated in cases of AD, but also can modulate AD pathogenesis. The following sections will discuss the critical roles and effects of Nurr1 in neurodegenerative diseases (Table 1). ...
Mechanisms of neuronal death in Alzheimer's disease
1
1996
... Neuronal death is the main pathogenic factor underlying neurodegenerative diseases such as AD and PD [67, 68]. Under neuropathological conditions, including neuroinflammation, excitotoxicity, and oxidative stress, neurons can be rescued by upregulation or activation of neuroprotective factors such as reactive oxygen species (ROS) scavengers or anti-apoptotic molecules [69-71]. Therefore, induction of neuroprotective factors may be a therapeutic strategy for the treatment of neuro-degeneration-related diseases. ...
Cell death pathways in Parkinson's disease: proximal triggers, distal effectors, and final steps
1
2009
... Neuronal death is the main pathogenic factor underlying neurodegenerative diseases such as AD and PD [67, 68]. Under neuropathological conditions, including neuroinflammation, excitotoxicity, and oxidative stress, neurons can be rescued by upregulation or activation of neuroprotective factors such as reactive oxygen species (ROS) scavengers or anti-apoptotic molecules [69-71]. Therefore, induction of neuroprotective factors may be a therapeutic strategy for the treatment of neuro-degeneration-related diseases. ...
How do adult neurons survive?
1
2005
... Neuronal death is the main pathogenic factor underlying neurodegenerative diseases such as AD and PD [67, 68]. Under neuropathological conditions, including neuroinflammation, excitotoxicity, and oxidative stress, neurons can be rescued by upregulation or activation of neuroprotective factors such as reactive oxygen species (ROS) scavengers or anti-apoptotic molecules [69-71]. Therefore, induction of neuroprotective factors may be a therapeutic strategy for the treatment of neuro-degeneration-related diseases. ...
Acteoside and its aglycones protect primary cultures of rat cortical cells from glutamate-induced excitotoxicity
0
2006
The antiapoptotic activity of melatonin in neurodegenerative diseases
1
2009
... Neuronal death is the main pathogenic factor underlying neurodegenerative diseases such as AD and PD [67, 68]. Under neuropathological conditions, including neuroinflammation, excitotoxicity, and oxidative stress, neurons can be rescued by upregulation or activation of neuroprotective factors such as reactive oxygen species (ROS) scavengers or anti-apoptotic molecules [69-71]. Therefore, induction of neuroprotective factors may be a therapeutic strategy for the treatment of neuro-degeneration-related diseases. ...
Nurr1 deficiency predisposes to lactacystin-induced dopaminergic neuron injury in vitro and in vivo
1
2008
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
A Nurr1 pathway for neuroprotection
0
2009
Conditional expression of Parkinson's disease-related mutant alpha-synuclein in the midbrain dopaminergic neurons causes progressive neurodegeneration and degradation of transcription factor nuclear receptor related 1
1
2012
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Nurr1 expression is modified by inflammation in microglia
1
2016
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Differential role of ERK in cAMP-induced Nurr1 expression in N2A and C6 cells
0
2004
Increased expression of the orphan nuclear receptor NURR1 in psoriasis and modulation following TNF-alpha inhibition
1
2008
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Dopamine Neuron Agenesis in Nurr1-Deficient Mice
3
1997
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
A Nurr1/CoREST pathway in microglia and astrocytes protects dopaminergic neurons from inflammation-induced death
9
2009
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... ]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Nuclear import and export signals control the subcellular localization of Nurr1 protein in response to oxidative stress
1
2013
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
NR4A orphan nuclear receptors as mediators of CREB-dependent neuroprotection
3
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... , 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Overexpression of midbrain-specific transcription factor Nurr1 modifies susceptibility of mouse neural stem cells to neurotoxins
0
2002
Generation of dopamine neurons with improved cell survival and phenotype maintenance using a degradation-resistant nurr1 mutant
3
2009
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
... . reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Covalent Modification and Regulation of the Nuclear Receptor Nurr1 by a Dopamine Metabolite
1
2019
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Implications of GABAergic Neurotransmission in Alzheimer's Disease
1
2016
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Loss of functional GABA(A) receptors in the Alzheimer diseased brain
1
2012
... Several studies have shown that Nurr1 has anti-apoptotic or neuroprotective roles against neuropathological stress or insults [56-60, 72-74]. Nurr1 expression is known to be regulated by various stimuli such as inflammatory cytokines, cAMP, and growth factors [75-77]. In comparison with WT mice, neurons from Nurr1 heterozygous mice exhibit greater vulnerability to neurotoxic challenges [59]. In addition, survival of dopaminergic neurons in the midbrain of mice was inhibited by genetic deletion of Nurr1 during development [78]. Moreover, it has been demonstrated that Nurr1 inhibits dopaminergic neuronal loss by suppressing inflammatory stimuli in the microglia and astrocytes [79]. Nurr1 changes its subcellular distribution in response to oxidative stress [80]. In addition, the ligand and agonist of Nurr1 showed neuroprotective effects on subsequent oxidative insult such as MPTP and 6-OHAD [17, 19, 20]. Especially, increased expression of Nurr1 by lentiviruses upregulated genes involved in ROS removal, such as Sesn3, Abl2, and Sod1, and demonstrated that Nurr1 is an essential mediator of CREB-dependent neuroprotection in oxidative stress [81]. Furthermore, Nurr1 overexpression protected neuronal stem cells against oxidative stress through downregulating cell death related protein such as caspase-3 and caspase-11 [60]. A number of studies have demonstrated the protective role and correlation of Nurr1 in cell death by oxidative stress, but the detailed molecular mechanism of Nurr1 against oxidative stress remains unclear [17, 19, 20, 60, 81-83]. Interestingly, a contemporary study reporting the direct binding of dopamine metabolite on Nurr1 and its stimulation of Nurr1 activity may provide an evidence for mechanism underlying the role of Nurr1 in sensing and responding the oxidative stress [84]. Moreover, Jo et al. reported that exogenous Nurr1 expression in neural precursor cells (NPCs) induced differentiation of dopaminergic neurons, higher resistance to toxic stimuli, and enhanced survival [83]. Although there has been only a few reports regarding the direct roles of Nurr1 in neuronal death in AD, interestingly, it has been demonstrated that Nurr1 is not only involved in the protection of dopaminergic neurons but also of GABA-positive neurons in vitro [79]. Since the levels of GABAergic neurotransmission as well as GABAergic signaling are significantly altered in AD [85, 86], it can be speculated that strategies for preserving GABAergic neurons by maintaining Nurr1 expression. All these reports suggest that Nurr1 may have neuroprotective effects against the pathogenesis of neurodegenerative diseases. A recent study has reported the protective role of Nurr1 in neuronal death in AD [16]. ...
Microglia: Housekeeper of the Central Nervous System
1
2018
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
Neuroinflammation in Alzheimer's disease
2
2015
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
Microglia-mediated neurotoxicity: uncovering the molecular mechanisms
1
2007
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
Ghrelin ameliorates cognitive dysfunction and neurodegeneration in intrahippocampal amyloid-beta1-42 oligomer-injected mice
4
2011
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... ]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
Bombycis excrementum reduces amyloid-beta oligomer-induced memory impairments, neurodegeneration, and neuroinflammation in mice
0
2014
6-Shogaol, an active constituent of ginger, attenuates neuroinflammation and cognitive deficits in animal models of dementia
2
2014
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... -92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
The NR4A subfamily of nuclear receptors: potential new therapeutic targets for the treatment of inflammatory diseases
2
2017
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
The nuclear orphan receptor Nr4a2 induces Foxp3 and regulates differentiation of CD4+ T cells
4
2011
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
... cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
Inflammation: a role for NR4A orphan nuclear receptors?
1
2011
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
Nurr1 expression and its modulation in microglia
2
2009
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
The N-terminal region of Nurr1 (a.a 1-31) is essential for its efficient degradation by the ubiquitin proteasome pathway
1
2013
... Neuroinflammation is one of the most important aspects of AD pathogenesis. Although resting glial cells maintain the microenvironment in the brain, activated glia contributes to neuronal damage by releasing neurotoxic molecules [87, 88]. Over-activated microglia and astrocytes release several detrimental compounds such as ROS, superoxide (O2•-), nitric oxide (•NO), and cytokines, which cause neuronal damage. The Aβ peptide is known to directly activate microglial cells, and activated cells are recruited around Aβ plaques before symptom development [89]. In addition, our previous studies revealed that Aβ oligomers (AβO) may induce both gliosis and neurodegeneration in the animal brain [90-92]. A significant increase in the number of microglial cells and decrease in the number of neurons were simultaneously observed in the brain of AβO-injected mice [90-92]. NR4A receptors are promptly activated by inflammatory stimuli, thus regulating not only initiation of inflammatory responses but also in the late stages of inflammation. NR4A receptors are involved in a negative feedback loop as modulators of inflammation [93]. Nurr1 can mediate inflammatory responses and regulate the function of immune cells [94, 95]. In microglia, inflammatory stimuli such as lipopolysaccharides (LPS) up-regulate Nurr1 mRNA expression [96]. Notably, Nurr1 shows potent anti-inflammatory activity in the CNS. In microglia and astrocytes, Nurr1 receptors inhibit the expression of pro-inflammatory cytokines, which are neurotoxic and eventually induce neuronal death, whereas reduction of Nurr1 enhances the inflammatory responses [79]. Nurr1 mediates the GSK3β-dependent repression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). Mechanistically, inflammatory signals induce Nurr1 phosphorylation and sumoylation, thus promoting Nurr1 binding to p65 and recruitment of the co-repressor for the RE1 silencing transcription factor (CoREST) complex to promoters of inflammatory genes, resulting in modulation of neuroinflammation [79]. Interestingly, a previous study revealed that deletion of amino acids 1-31 from the N-terminal region of Nurr1 yields better performance in transcription compared to full-length Nurr1 [97]. In addition, a recent study has provided direct evidence that modulation of Nurr1 can be involved in Aβ-mediated neuroinflammation [16]. In conclusion, Nurr1 can serve as a possible therapeutic target for treatment of AD by inhibiting the transcription of inflammatory genes and modulating the function of immune cells. ...
Inflammation and Alzheimer's disease
1
2000
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
The blood-brain barrier and cerebrovascular pathology in Alzheimer's disease
1
1999
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
Occurrence of T cells in the brain of Alzheimer's disease and other neurological diseases
1
2002
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
Increased activity of Th-17 and Th-9 lymphocytes and a skewing of the post-thymic differentiation pathway are seen in Alzheimer's disease
1
2011
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
Hypoxia enhances stimulating effect of amyloid beta peptide (25-35) for interleukin 17 and T helper lymphocyte subtype 17 upregulation in cultured peripheral blood mononuclear cells
0
2009
IFN-gamma Production by amyloid beta-specific Th1 cells promotes microglial activation and increases plaque burden in a mouse model of Alzheimer's disease
0
2013
Th17 cell-mediated neuroinflammation is involved in neurodegeneration of abeta1-42-induced Alzheimer's disease model rats
1
2013
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
T cells specifically targeted to amyloid plaques enhance plaque clearance in a mouse model of Alzheimer's disease
1
2010
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
DNA immunization against amyloid beta 42 has high potential as safe therapy for Alzheimer's disease as it diminishes antigen-specific Th1 and Th17 cell proliferation
1
2011
... The early stage of AD involves both the activation of microglia and astrocytes overexpressing cytokines around the Aβ plaques and an increase in the levels of pro-inflammatory cytokines in the peripheral blood [98]. In addition, the cross-talk between the peripheral blood and the brain via a damaged blood-brain barrier (BBB) may be enhanced in AD patients, thereby contributing to neuroinflammation in AD [99]. As evidenced, an increase in the number of T cells was observed in the brain parenchyma of AD patients [100]. Several lines of evidence have indicated that modulation of T helper (Th) cells may be involved in AD pathologies [101-104]. In particular, CD4 and CD8 T cells specifically migrated to the Aβ plaques, thus enhancing elimination of Aβ plaques [105]. Moreover, immunization with the Aβ42 DNA trimmer was shown to suppress antigen-specific Th17 and Th1 cell proliferation [106]. ...
Monocytes and CD4+ T cells contribution to the under-expression of NR4A2 and TNFAIP3 genes in patients with multiple sclerosis
1
2014
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Learning from nature: pregnancy changes the expression of inflammation-related genes in patients with multiple sclerosis
1
2010
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Microarray analysis identifies altered regulation of nuclear receptor family members in the pre-disease state of multiple sclerosis
1
2010
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
T cell gene expression profiling identifies distinct subgroups of Japanese multiple sclerosis patients
1
2006
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Live imaging of effector cell trafficking and autoantigen recognition within the unfolding autoimmune encephalomyelitis lesion
1
2005
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
Towards understanding the role of orphan nuclear receptor NR4A2 in Th17 cell-mediated central nervous system autoimmunity: An experimental approach using an animal model of multiple sclerosis
1
2014
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Nurr1 reduction influences the onset of chronic EAE in mice
1
2015
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Modulation of orphan nuclear receptor NURR1 expression by methotrexate in human inflammatory joint disease involves adenosine A2A receptor-mediated responses
1
2005
... Peripheral blood mononuclear cells, CD4+ T cells and monocytes obtained from patients with multiple sclerosis (MS) have been reported to have decreased Nurr1 gene expression [107, 108]. In addition, microarray analysis showed that key nuclear receptor family genes such as NR4A1 (Nur77) and NR4A2 (Nurr1), which are important for the nuclear receptor-dependent apoptosis in the peripheral blood of the pre-disease state in MS patients, were suppressed [109]. In contrast, over-expression of Nurr1 was observed in peripheral blood T cells derived from relapsing-remitting MS patients [110]. In an experimental autoimmune encephalomyelitis (EAE) model that serves as an animal model of MS, effector T cells infiltrated the parenchyma of the CNS [111]. Nurr1 is selectively over-expressed in T cells in the peripheral blood whereas expression of Nurr1 in the T cells in lymphoid organs did change during the induction of EAE. In addition, interleukin (IL)-17-producing tyrosine hydroxylase (TH)-positive cells express Nurr1 regardless of interferon (IFN)-γ secretion [112]. Hence, since Nurr1 could be a useful biomarker for determining the status of T cells in MS [113], assessment of Nurr1 expression in T cells in AD could also be useful to identify changes in T cell activation status. In addition, heterozygous Nurr1 mice promoted early onset of EAE and increased the infiltration of inflammatory cells into the spinal cord [114], indicating that Nurr1 is involved in the pathophysiology of autoimmune diseases such MS. Consequently, Nurr1 could be an innovative therapeutic target for various autoimmune diseases. In human inflammatory joint disease, Nurr1 has been identified as a molecular target of methotrexate (MTX)-related reactions. MTX considerably subdues Nurr1 expression in patients with active psoriatic arthritis. In the synovial tissue, MTX selectively regulates Nurr1 induced by inflammatory stimulation and also modulates expression of growth factors in resident cells. Moreover, suppressive effect of MTX on Nurr1 expression is mediated by adenosine release [115]. ...
Functional redundancy of the Nur77 and Nor-1 orphan steroid receptors in T-cell apoptosis
1
1997
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
T cell receptor signal strength in Treg and iNKT cell development demonstrated by a novel fluorescent reporter mouse
1
2011
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis
2
2013
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
The adaptive immune system restrains Alzheimer's disease pathogenesis by modulating microglial function
1
2016
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
Role of the peripheral innate immune system in the development of Alzheimer's disease
1
2018
... Because Nr4a receptors play an important role in initiating regulatory T (Treg) cell development in the thymus [94, 116, 117], the role of Nr4a receptors in peripheral immune regulation has been examined in Nr4a-triple-knockout (Nr4a-TKO) mice, Nur77 (Nr4a1)-/-, Nurr1 (Nr4a2)-/-, and Nor-1 (Nr4a3)-/-. The Nr4a-TKO mice could not produce Treg cells and died early due to systemic autoimmunity [118]. Specifically, Nurr1 binds directly to the Foxp3 promoter, leading to activation of transcription and the development of Treg cells. In addition, Nurr1 has been reported to bind directly to the regulatory regions of Foxp3, at which Nurr1 intervenes via histone modifications. Furthermore, in Nurr1-deficient T cells, aberrant Th1 induction is increased but Treg cell induction is rather decreased [94]. In conclusion, Nurr1 plays central roles not only in regulating the induction and suppressive functions of Treg cells but also in inhibiting aberrant Th1 induction. Moreover, Nurr1 can regulate the Th17 cell-mediated autoimmune inflammation, contributing to the pathogenesis of MS, an immune disease of the nervous system [112]. Since peripheral immune functions are involved in the pathogenesis of AD [119, 120], modulation of peripheral immune responses though Nurr1 may be a potential therapeutic strategy against AD. ...
Aberrant expression of mitotic cdc2/cyclin B1 kinase in degenerating neurons of Alzheimer's disease brain
2
1997
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
... complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Neuronal cell cycle: the neuron itself and its circumstances
1
2015
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
DNA replication precedes neuronal cell death in Alzheimer's disease
1
2001
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Neuronal cell death is preceded by cell cycle events at all stages of Alzheimer's disease
1
2003
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Neuronal cell cycle re-entry markers are altered in the senescence accelerated mouse P8 (SAMP8)
1
2012
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Conversion of p35 to p25 deregulates Cdk5 activity and promotes neurodegeneration
1
1999
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Amyloid-beta signals through tau to drive ectopic neuronal cell cycle re-entry in Alzheimer's disease
1
2013
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Neuron-specific phosphorylation of Alzheimer's beta-amyloid precursor protein by cyclin-dependent kinase 5
1
2000
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Regulation of amyloid precursor protein (APP) phosphorylation and processing by p35/Cdk5 and p25/Cdk5
0
2003
Cell cycle-dependent regulation of the phosphorylation and metabolism of the Alzheimer amyloid precursor protein
1
1994
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
APP processing is regulated by cytoplasmic phosphorylation
1
2003
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
U1 snRNP Alteration and Neuronal Cell Cycle Reentry in Alzheimer Disease
1
2018
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009
1
2009
... Neurons are generally considered as postmitotic cells, and can cell undergo cell cycle re-entry in neurodegenerative conditions [121, 122]. Basal forebrain and hippocampal pyramid neurons in the brain with AD have been reported to progress from the G1-phase to the S-phase [123]. In brains with AD and mild cognitive impairment, the expression levels of markers associated with cell cycle and proliferation, such as the proliferating cell nuclear antigen (PCNA), cyclin D, and B1, are increased in various regions including the entorhinal cortex, hippocampus, and nucleus basalis of Meynert [124]. Moreover, the presence of active cdc2 and cyclin B1 complex was observed in the AD brain tissue [121]. In addition, senescence-accelerated mice-prone 8, which show the major pathologic features of AD such as Aβ accumulation and tau phosphorylation, not only show enhanced CDK5 and GSK3β expression, but also show increased expression of various cell-cycle re-entry markers such as CDK2, cyclins A, D1, E, and B [125]. There are several reports on the correlation between cell-cycle-related kinases and histological hallmarks of AD. p25, a truncated form of the subunit p35 that activates CDK5, is not readily degraded and is found to accumulate in the brain of patients with AD. Subsequently, the formation of the p25/CDK5 complex induced tau hyperphosphorylation and apoptosis [126]. In addition, soluble Aβ oligomers also promoted neuronal cell-cycle re-entry via phosphorylation of tau [127]; cell-cycle progression through CDK5 and CDC2 kinases induced phosphorylation of the amyloid precursor protein (APP) [128-130], and phosphorylation of APP facilitated Aβ generation [131]. Moreover, a broad promotion of the cell cycle in the AD brain leads to a mitotic catastrophe, which is the result of dysregulated or failed mitosis, suggesting that this may be one of the mechanisms of neuronal death in AD [132, 133]. ...
Induction of cell cycle arrest and morphological differentiation by Nurr1 and retinoids in dopamine MN9D cells
2
2001
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
Nuclear receptor Nurr1 is expressed in and is associated with human restenosis and inhibits vascular lesion formation in mice involving inhibition of smooth muscle cell proliferation and inflammation
4
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
... In human atherosclerosis, expression of NR4A including Nurr1 receptors is increased in macrophages after inflammatory stimulation [177]. In addition, overexpression of Nurr1 inhibited vascular lesion formation through reduction of smooth muscle cell proliferation and inflammatory response [135, 178]. These findings prove that endogenous Nurr1 suppresses macrophage activation, foam-cell formation, and further differentiation. Thus, it provides further evidence that atherogenesis could be prevented by modulation of Nurr1 expression. Therefore, Nurr1 may be proposed as a novel therapeutic target for preventing vascular disruption-related diseases mediated by AD. ...
The orphan nuclear receptor Nurr1 restricts the proliferation of haematopoietic stem cells
2
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
Nuclear receptors Nur77 and Nurr1 modulate mesenchymal stromal cell migration
1
2012
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
Nurr1 blocks the mitogenic effect of FGF-2 and EGF, inducing olfactory bulb neural stem cells to adopt dopaminergic and dopaminergic-GABAergic neuronal phenotypes
1
2015
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
Nurr1 promotes intestinal regeneration after ischemia/reperfusion injury by inhibiting the expression of p21 (Waf1/Cip1)
2
2017
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... The role of Nurr1 in the cell cycle has been suggested after assessment of cell-cycle-related molecules in various cells. In the dopamine-synthesizing cell line (MN9D cells), Nurr1 promoted cell-cycle arrest in the G1 phase as well as morphological differentiation, and these effects did not require the formation of heterodimers with retinoid X receptors (RXR) [134]. In vascular smooth muscle cells, lentivirus-mediated Nurr1 overexpression inhibited proliferation with increased expression of the crucial cell-cycle inhibitor p27Kip1 which induces G1 cell-cycle arrest [135]. Similarly, in hematopoietic stem cells, Nurr1 overexpression restricted cell proliferation by upregulating the expression of p18, which inhibits the cyclin D/CDK4/6 complexes required for cell-cycle progression in G1-phase [136]. In addition, the mechanism by which Nurr1 promotes migration and inhibits proliferation in mesenchymal stem cells (MSCs) may involve the ability of Nurr1 to reduce the percentage of cells in the S-phase [137]. Furthermore, overexpression of Nurr1 in olfactory bulb stem cells induces cell-cycle exit, inhibits proliferation, and induces a TH neuronal fate mediated by Fgfr2 expression [138]. In contrast, Nurr1 induced after intestinal ischemia/reperfusion injury promoted proliferation of intestinal epithelial cells via inhibition of p21Waf1/cIP1 gene transcription [139]. Moreover, in the mouse hippocampus and adult hippocampal neural precursor cells, pharmacological stimulation of Nurr1 with a Nurr1 agonist resulted in increased proliferation as well as phosphorylation of Akt and Erk1/2 [18]. ...
Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior
2
2016
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
... ]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
Mammalian neural stem cells
1
2000
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
Neurogenesis in the dentate gyrus: carrying the message or dictating the tone
1
2013
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory?
1
2010
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
Adult hippocampal neurogenesis and its role in Alzheimer's disease
2
2011
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
... ]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
Neurogenesis and Alzheimer's disease: At the crossroads
0
2010
Amyloid-beta accumulation, neurogenesis, behavior, and the age of rats
0
2014
The amyloid precursor protein controls adult hippocampal neurogenesis through GABAergic interneurons
0
2014
Amyloid precursor protein regulates neurogenesis by antagonizing miR-574-5p in the developing cerebral cortex
1
2014
... Neurons are generated and differentiated from neural stem cells in the adult brain. This process is called adult neurogenesis, and takes place mainly in two brain regions, the subgranular zone of the hippocampal dentate gyrus (SGZ) and the subventricular zone of the lateral ventricle (SVZ) [140, 141]. In particular, adult hippocampal neurogenesis at SGZ regulates learning and memory functions by generating newborn neurons derived from neural stem cells [140, 142, 143]. Studies have shown that altered hippocampal neurogenesis occurs in the early stage of AD even prior to pathologic changes [144]. Several key molecular players involved in AD pathogenesis have been found to regulate hippocampal neurogenesis [144-148]. ...
Generation of functional dopamine neurons from neural precursor cells isolated from the subventricular zone and white matter of the adult rat brain using Nurr1 overexpression
3
2007
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
Selective Agenesis of Mesencephalic Dopaminergic Neurons in Nurr1-Deficient Mice
2
1999
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
Dopamine neurons derived from embryonic stem cells function in an animal model of Parkinson's disease
1
2002
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
Orphan nuclear receptor Nurr1 induces neuron differentiation from embryonic cortical precursor cells via an extrinsic paracrine mechanism
1
2009
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
Transplantation of post-mitotic human neuroteratocarcinoma-overexpressing Nurr1 cells provides therapeutic benefits in experimental stroke: in vitro evidence of expedited neuronal differentiation and GDNF secretion
1
2007
... A recent study has shown that Nurr1 expression was dynamic following acute ischemia induced by middle cerebral artery occlusion (MCAO)/reperfusion in a rat model. There was a negative correlation between Nurr1 and infarct volume up to 12 hours after ligation, but a positive correlation was observed after 24 hours. Nurr1 overexpression inhibited tumor necrosis factor-α (TNF-α) levels in microglia. Increase of Nurr1 expression through suppression of miR-145-5p, a negative regulator of Nurr1 alleviated infarct volume and improved the neurological outcomes in an acute stroke model [223]. In addition, transplantation of Nurr1-overexpressing human embryonal carcinoma cells into the ischemic striatum restored the behavioral disorder in a transient MCAO rat model [153]. Considering that TNF-α released from Aβ-activated microglia is a key cytokine causing cell cycle events, which are related to pathogenesis of neuronal death in AD [224], these regulatory effects of Nurr1 on microglial activation could be a therapeutic target for AD as well as ischemic stroke. ...
Proneural bHLH neurogenin 2 differentially regulates Nurr1-induced dopamine neuron differentiation in rat and mouse neural precursor cells in vitro
0
2008
Nurr1 promotes neurogenesis of dopaminergic neuron and represses inflammatory factors in the transwell coculture system of neural stem cells and microglia
2
2018
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
Age-dependent decrease of Nurr1 protein expression in the gerbil hippocampus
1
2018
... A recent study demonstrated that treatment with the Nurr1 agonist amodiaquine (AQ) in mice significantly contributed to enhanced adult hippocampal neurogenesis, resulting in enhancement of cognitive function. Moreover, knockdown of Nurr1 inhibited proliferation of adult hippocampal neural stem cells [18]. In addition, overexpression of Nurr1 in NPCs isolated from the SVZ of adult rats resulted in functional dopaminergic neurons. Transplantation of Nurr1-induced dopaminergic neurons lead to differentiation and integration in vivo, and improved the behavioral disorders of parkinsonian rats [149]. Furthermore, in vivo studies showed that Nurr1-deficient mice exhibited deficits in the differentiation of dopaminergic neurons in the ventral midbrain [78, 150]. In support of this notion, Nurr1 is not only well known to play a key role in the differentiation and maturation of dopaminergic neurons [151-155] but may also exert an important role in neurogenesis [156]. These studies suggest that Nurr1 may contribute to the rescue of impaired adult neurogenesis in AD. Indeed, a recent study has shown that administration of the Nurr1 agonist AQ can reverse impaired neuronal fate specification of hippocampal neural stem cells in Aβ-overexpressing mice [16]. ...
The role of amyloid-beta in the regulation of memory
1
2014
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
A learning deficit related to age and beta-amyloid plaques in a mouse model of Alzheimer's disease
0
2000
Adult neurogenesis: a potential tool for early diagnosis in Alzheimer's disease?
1
2010
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
NMDA neurotransmission dysfunction in mild cognitive impairment and Alzheimer's disease
1
2014
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
Dysfunction of NMDA receptors in Alzheimer's disease
1
2016
... Aβ is known to be a major contributor of memory impairment in AD, and results in cognitive deficits by inducing neuroinflammation, neuronal death, inhibition of synaptic transmission, synaptic loss, and impairment of adult neurogenesis [88, 157-159]. Furthermore, under AD conditions, there are other pathways that cause cognitive dysfunction and memory decline, such as abnormal activity of the NMDA receptor [160, 161]. Recent studies have suggested that Nurr1 may play a role in the regulation of cognitive function, since hippocampus-dependent memories were impaired in Nurr1 knockdown or NR4A dominant-negative transgenic mice [48, 49, 54] and an increase in Nurr1 mRNA expression was observed when mice were submitted to spatial learning tasks [50]. Thus, it is important to investigate whether Nurr1 has a direct influence on AD-related cognitive functions. ...
CREB, synapses and memory disorders: Past progress and future challenges
1
2005
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
CREB has a context-dependent role in activity-regulated transcription and maintains neuronal cholesterol homeostasis
1
2008
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory
1
1997
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Recruitment of long-lasting and protein kinase A-dependent long-term potentiation in the CA1 region of hippocampus requires repeated tetanization
0
1994
Two time periods of hippocampal mRNA synthesis are required for memory consolidation of fear-motivated learning
0
2002
Requirement of a critical period of transcription for induction of a late phase of LTP
0
1994
Reversible inhibition of CREB/ATF transcription factors in region CA1 of the dorsal hippocampus disrupts hippocampus-dependent spatial memory
2
2002
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Rapidly induced gene networks following induction of long-term potentiation at perforant path synapses in vivo
1
2011
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Sequential steps underlying neuronal plasticity induced by a transient exposure to gabazine
1
2010
... Several studies have discovered that Nurr1 regulates learning and memory functions [15, 49-55]. In many systems, the CREB signaling pathway is important for transcription of memory-related genes [162], and this pathway controls transcription of Nurr1 [163]. Additionally, long-term memory in the hippocampus depends on CREB-related pathways that can regulate Nurr1 expression [48, 164-168]. Several studies demonstrated that behavioral task training increases Nurr1 gene expression in subregions of the hippocampus [50, 53, 54]. Moreover, a network analysis of genes in the dentate gyri of long-term potentiation-induced rats revealed that expression of NR4A nuclear receptors, including Nurr1, was upregulated [169]. Expression of Nurr1 in cultured hippocampal neurons is revealed to be increased after treatment with GABA antagonists [170]. ...
Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB: CBP-dependent transcriptional activation
2
2007
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
Effect of intermittent preventive treatment of malaria on health and education in schoolchildren: a cluster-randomised, double-blind, placebo-controlled trial
1
2008
... Inhibition of histone deacetylase (HDAC) by trichostatin A increases Nurr1 expression, and enhances memory [171]. Similarly, enhancement of memory by HDAC inhibition is attenuated and memory enhancement is impaired by negative protein expression of Nr4a family receptors [54]. Moreover, memory functions are impaired by siRNA knockdown of Nr4a2 [53] and generation of heterozygosity for the Nurr1 gene [15]. A recent study also revealed the importance of the NR4A family, including Nurr1, showing that hippocampal long-term potentiation was impaired in dominant-negative Nr4a transgenic mice [55]. Interestingly, although the precise mechanism for cognitive enhancement is unclear, one placebo-controlled study showed that the intermittent preventive administration of AQ, an antimalarial agent and Nurr1 agonist, enhanced cognitive performance in semi-immune schoolchildren [172]. Furthermore, a recent study has demonstrated that administration of the Nurr1 agonist AQ restored damaged spatial working memory in Aβ-overexpressing mice [16]. However, how Nurr1 affects AD patients with memory failure has not been examined, and we anticipate that further studies on Nurr1 will uncover its importance on cognitive functions in brains with AD. ...
Cerebral amyloid angiopathy: a systematic review
1
2011
... Cerebral amyloid angiopathy (CAA) is an AD-related histopathology showing pathological changes such as deposits of Aβ in the blood vessels of the CNS and the walls of leptomeningeal arteries [173]. CAA is not only associated with the Aβ burden of the brain parenchyma, but also occasionally induces necrosis resulting in cerebral hemorrhage [174, 175]. However, the degree of CAA varies between AD brains, and the majority of AD patients showing microvascular amyloid deposits do not experience cerebral hemorrhage [176]. Nonetheless, because CAA could contribute to cortical dysfunction, regulation of vascular Aβ accumulation is suggested for mitigation of secondary AD pathology. ...
Cerebral amyloid angiopathy and thrombolysis-related intracerebral haemorrhage
1
2004
... Cerebral amyloid angiopathy (CAA) is an AD-related histopathology showing pathological changes such as deposits of Aβ in the blood vessels of the CNS and the walls of leptomeningeal arteries [173]. CAA is not only associated with the Aβ burden of the brain parenchyma, but also occasionally induces necrosis resulting in cerebral hemorrhage [174, 175]. However, the degree of CAA varies between AD brains, and the majority of AD patients showing microvascular amyloid deposits do not experience cerebral hemorrhage [176]. Nonetheless, because CAA could contribute to cortical dysfunction, regulation of vascular Aβ accumulation is suggested for mitigation of secondary AD pathology. ...
Review: sporadic cerebral amyloid angiopathy
1
2011
... Cerebral amyloid angiopathy (CAA) is an AD-related histopathology showing pathological changes such as deposits of Aβ in the blood vessels of the CNS and the walls of leptomeningeal arteries [173]. CAA is not only associated with the Aβ burden of the brain parenchyma, but also occasionally induces necrosis resulting in cerebral hemorrhage [174, 175]. However, the degree of CAA varies between AD brains, and the majority of AD patients showing microvascular amyloid deposits do not experience cerebral hemorrhage [176]. Nonetheless, because CAA could contribute to cortical dysfunction, regulation of vascular Aβ accumulation is suggested for mitigation of secondary AD pathology. ...
Alzheimer's disease: genes, proteins, and therapy
1
2001
... Cerebral amyloid angiopathy (CAA) is an AD-related histopathology showing pathological changes such as deposits of Aβ in the blood vessels of the CNS and the walls of leptomeningeal arteries [173]. CAA is not only associated with the Aβ burden of the brain parenchyma, but also occasionally induces necrosis resulting in cerebral hemorrhage [174, 175]. However, the degree of CAA varies between AD brains, and the majority of AD patients showing microvascular amyloid deposits do not experience cerebral hemorrhage [176]. Nonetheless, because CAA could contribute to cortical dysfunction, regulation of vascular Aβ accumulation is suggested for mitigation of secondary AD pathology. ...
Induction of NR4A orphan nuclear receptor expression in macrophages in response to inflammatory stimuli
1
2005
... In human atherosclerosis, expression of NR4A including Nurr1 receptors is increased in macrophages after inflammatory stimulation [177]. In addition, overexpression of Nurr1 inhibited vascular lesion formation through reduction of smooth muscle cell proliferation and inflammatory response [135, 178]. These findings prove that endogenous Nurr1 suppresses macrophage activation, foam-cell formation, and further differentiation. Thus, it provides further evidence that atherogenesis could be prevented by modulation of Nurr1 expression. Therefore, Nurr1 may be proposed as a novel therapeutic target for preventing vascular disruption-related diseases mediated by AD. ...
Nuclear receptors Nur77, Nurr1, and NOR-1 expressed in atherosclerotic lesion macrophages reduce lipid loading and inflammatory responses
2
2006
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... In human atherosclerosis, expression of NR4A including Nurr1 receptors is increased in macrophages after inflammatory stimulation [177]. In addition, overexpression of Nurr1 inhibited vascular lesion formation through reduction of smooth muscle cell proliferation and inflammatory response [135, 178]. These findings prove that endogenous Nurr1 suppresses macrophage activation, foam-cell formation, and further differentiation. Thus, it provides further evidence that atherogenesis could be prevented by modulation of Nurr1 expression. Therefore, Nurr1 may be proposed as a novel therapeutic target for preventing vascular disruption-related diseases mediated by AD. ...
Increased tau phosphorylation and cleavage in mouse models of type 1 and type 2 diabetes
1
2009
... There are a number of studies describing both a correlation between type 2 diabetes and the risk of AD, and an association of hyperglycemia with AD pathophysiology [179, 180]. In addition, it has been reported that excess weight in middle age is related to increased risk of AD [181]. Metabolic hormones such as insulin, leptin, ghrelin, and adiponectin have been reported to have therapeutic potential and are also involved in the pathogenesis of AD [182]. Since AD could be accompanied by metabolic disturbances, normalization of metabolism will provide new insights into AD treatment. ...
Diabetes mellitus and the risk of dementia: The Rotterdam Study
1
1999
... There are a number of studies describing both a correlation between type 2 diabetes and the risk of AD, and an association of hyperglycemia with AD pathophysiology [179, 180]. In addition, it has been reported that excess weight in middle age is related to increased risk of AD [181]. Metabolic hormones such as insulin, leptin, ghrelin, and adiponectin have been reported to have therapeutic potential and are also involved in the pathogenesis of AD [182]. Since AD could be accompanied by metabolic disturbances, normalization of metabolism will provide new insights into AD treatment. ...
Body mass index in midlife and risk of Alzheimer disease and vascular dementia
1
2007
... There are a number of studies describing both a correlation between type 2 diabetes and the risk of AD, and an association of hyperglycemia with AD pathophysiology [179, 180]. In addition, it has been reported that excess weight in middle age is related to increased risk of AD [181]. Metabolic hormones such as insulin, leptin, ghrelin, and adiponectin have been reported to have therapeutic potential and are also involved in the pathogenesis of AD [182]. Since AD could be accompanied by metabolic disturbances, normalization of metabolism will provide new insights into AD treatment. ...
Metabolic dysfunction in Alzheimer's disease and related neurodegenerative disorders
1
2012
... There are a number of studies describing both a correlation between type 2 diabetes and the risk of AD, and an association of hyperglycemia with AD pathophysiology [179, 180]. In addition, it has been reported that excess weight in middle age is related to increased risk of AD [181]. Metabolic hormones such as insulin, leptin, ghrelin, and adiponectin have been reported to have therapeutic potential and are also involved in the pathogenesis of AD [182]. Since AD could be accompanied by metabolic disturbances, normalization of metabolism will provide new insights into AD treatment. ...
NR4A orphan nuclear receptors: transcriptional regulators of gene expression in metabolism and vascular biology
2
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... There are several studies showing the roles of Nr4a receptors associated with metabolic functions. In particular, it has been reported that aberrant expression of Nurr1 is correlated to glucose metabolism and metabolic syndrome [183, 184]. Moreover, Nr4a receptors are expressed under metabolism-related stimuli including cold, fatty acids, glucose, insulin, and cholesterol. Therefore, it has been suggested that Nr4a receptors can be therapeutic targets for metabolism-related disorders [185]. There is also compelling evidence suggesting that Nr4a receptors show a potent association with the course of type 2 diabetes through regulation of insulin sensitivity and glucose homeostasis. Even though the mechanisms underlying the regulation of glucose metabolism by Nr4a receptors have not been elucidated, Nr4a receptors are identified as potential biological targets for diabetic patients [186]. However, the association of Nurr1 with carbohydrate metabolism is currently unclear. Therefore, identification of the correlation between Nurr1 and metabolic disorders in AD will be a novel pioneering field of study. ...
Decreased expression levels of Nurr1 are associated with chronic inflammation in patients with type 2 diabetes
2
2015
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... There are several studies showing the roles of Nr4a receptors associated with metabolic functions. In particular, it has been reported that aberrant expression of Nurr1 is correlated to glucose metabolism and metabolic syndrome [183, 184]. Moreover, Nr4a receptors are expressed under metabolism-related stimuli including cold, fatty acids, glucose, insulin, and cholesterol. Therefore, it has been suggested that Nr4a receptors can be therapeutic targets for metabolism-related disorders [185]. There is also compelling evidence suggesting that Nr4a receptors show a potent association with the course of type 2 diabetes through regulation of insulin sensitivity and glucose homeostasis. Even though the mechanisms underlying the regulation of glucose metabolism by Nr4a receptors have not been elucidated, Nr4a receptors are identified as potential biological targets for diabetic patients [186]. However, the association of Nurr1 with carbohydrate metabolism is currently unclear. Therefore, identification of the correlation between Nurr1 and metabolic disorders in AD will be a novel pioneering field of study. ...
Minireview: Nuclear hormone receptor 4A signaling: implications for metabolic disease
2
2010
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... There are several studies showing the roles of Nr4a receptors associated with metabolic functions. In particular, it has been reported that aberrant expression of Nurr1 is correlated to glucose metabolism and metabolic syndrome [183, 184]. Moreover, Nr4a receptors are expressed under metabolism-related stimuli including cold, fatty acids, glucose, insulin, and cholesterol. Therefore, it has been suggested that Nr4a receptors can be therapeutic targets for metabolism-related disorders [185]. There is also compelling evidence suggesting that Nr4a receptors show a potent association with the course of type 2 diabetes through regulation of insulin sensitivity and glucose homeostasis. Even though the mechanisms underlying the regulation of glucose metabolism by Nr4a receptors have not been elucidated, Nr4a receptors are identified as potential biological targets for diabetic patients [186]. However, the association of Nurr1 with carbohydrate metabolism is currently unclear. Therefore, identification of the correlation between Nurr1 and metabolic disorders in AD will be a novel pioneering field of study. ...
NR4A orphan nuclear receptors in glucose homeostasis: a minireview
2
2013
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... There are several studies showing the roles of Nr4a receptors associated with metabolic functions. In particular, it has been reported that aberrant expression of Nurr1 is correlated to glucose metabolism and metabolic syndrome [183, 184]. Moreover, Nr4a receptors are expressed under metabolism-related stimuli including cold, fatty acids, glucose, insulin, and cholesterol. Therefore, it has been suggested that Nr4a receptors can be therapeutic targets for metabolism-related disorders [185]. There is also compelling evidence suggesting that Nr4a receptors show a potent association with the course of type 2 diabetes through regulation of insulin sensitivity and glucose homeostasis. Even though the mechanisms underlying the regulation of glucose metabolism by Nr4a receptors have not been elucidated, Nr4a receptors are identified as potential biological targets for diabetic patients [186]. However, the association of Nurr1 with carbohydrate metabolism is currently unclear. Therefore, identification of the correlation between Nurr1 and metabolic disorders in AD will be a novel pioneering field of study. ...
The dorsal diencephalic conduction system: a review of the anatomy and functions of the habenular complex
1
1982
... The habenula is a part of the epithalamus in the diencephalon, located dorsal-medial to the posterior thalamus. It is divided into two regions: (1) the lateral habenula (LHb), which is innervated by the rostromedial tegmental nucleus (RMTg) to the dopamine-related regions, such as the ventral tegmental area (VTA) and substantia nigra (SN) or the serotonin system, such as the dorsal raphe nucleus and median raphe nucleus, and (2) the medial habenula (MHb), which is innervated with the serotonin system through the interpeduncular nucleus (IPN) [187-190]. The habenula, which is connected to the limbic system, basal ganglia, and pineal gland, is involved in the reward system and cognitive functions such as learning, memory, and attention [191]. Therefore, it has been suggested that the habenula may be involved in psychiatric disorders such as depression, schizophrenia, and drug-induced psychosis [192]. ...
Cytoarchitecture, fiber connections, and some histochemical aspects of the interpeduncular nucleus in the rat
0
1986
Efferent connections of the habenular nuclei in the rat
0
1979
The mesopontine rostromedial tegmental nucleus: A structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta
1
2009
... The habenula is a part of the epithalamus in the diencephalon, located dorsal-medial to the posterior thalamus. It is divided into two regions: (1) the lateral habenula (LHb), which is innervated by the rostromedial tegmental nucleus (RMTg) to the dopamine-related regions, such as the ventral tegmental area (VTA) and substantia nigra (SN) or the serotonin system, such as the dorsal raphe nucleus and median raphe nucleus, and (2) the medial habenula (MHb), which is innervated with the serotonin system through the interpeduncular nucleus (IPN) [187-190]. The habenula, which is connected to the limbic system, basal ganglia, and pineal gland, is involved in the reward system and cognitive functions such as learning, memory, and attention [191]. Therefore, it has been suggested that the habenula may be involved in psychiatric disorders such as depression, schizophrenia, and drug-induced psychosis [192]. ...
A conductor hidden in the orchestra? Role of the habenular complex in monoamine transmission and cognition
2
2007
... The habenula is a part of the epithalamus in the diencephalon, located dorsal-medial to the posterior thalamus. It is divided into two regions: (1) the lateral habenula (LHb), which is innervated by the rostromedial tegmental nucleus (RMTg) to the dopamine-related regions, such as the ventral tegmental area (VTA) and substantia nigra (SN) or the serotonin system, such as the dorsal raphe nucleus and median raphe nucleus, and (2) the medial habenula (MHb), which is innervated with the serotonin system through the interpeduncular nucleus (IPN) [187-190]. The habenula, which is connected to the limbic system, basal ganglia, and pineal gland, is involved in the reward system and cognitive functions such as learning, memory, and attention [191]. Therefore, it has been suggested that the habenula may be involved in psychiatric disorders such as depression, schizophrenia, and drug-induced psychosis [192]. ...
... In adults and during development, the Nurr1 and Nr4a2 genes are robustly and specifically expressed in the MHb expressing both the choline acetyltransferase in the ventral part and the neuropeptide SP in the dorsal part [191, 193-195]. In addition, Nurr1 has been reported to mediate a gene pathway involved in habenula development regulated by the POU-domain transcription factor Brn3a [195]. Notably, several studies have suggested that habenular activity is involved in depression, which is one of the most common psychiatric symptoms in AD and a risk factor for AD development. ...
The habenula: from stress evasion to value-based decision-making
1
2010
... The habenula is a part of the epithalamus in the diencephalon, located dorsal-medial to the posterior thalamus. It is divided into two regions: (1) the lateral habenula (LHb), which is innervated by the rostromedial tegmental nucleus (RMTg) to the dopamine-related regions, such as the ventral tegmental area (VTA) and substantia nigra (SN) or the serotonin system, such as the dorsal raphe nucleus and median raphe nucleus, and (2) the medial habenula (MHb), which is innervated with the serotonin system through the interpeduncular nucleus (IPN) [187-190]. The habenula, which is connected to the limbic system, basal ganglia, and pineal gland, is involved in the reward system and cognitive functions such as learning, memory, and attention [191]. Therefore, it has been suggested that the habenula may be involved in psychiatric disorders such as depression, schizophrenia, and drug-induced psychosis [192]. ...
Topography of cholinergic and substance P pathways in the habenulo-interpeduncular system of the rat. An immunocytochemical and microchemical approach
1
1987
... In adults and during development, the Nurr1 and Nr4a2 genes are robustly and specifically expressed in the MHb expressing both the choline acetyltransferase in the ventral part and the neuropeptide SP in the dorsal part [191, 193-195]. In addition, Nurr1 has been reported to mediate a gene pathway involved in habenula development regulated by the POU-domain transcription factor Brn3a [195]. Notably, several studies have suggested that habenular activity is involved in depression, which is one of the most common psychiatric symptoms in AD and a risk factor for AD development. ...
Genome-wide atlas of gene expression in the adult mouse brain
0
2007
Brn3a and Nurr1 mediate a gene regulatory pathway for habenula development
2
2009
... In adults and during development, the Nurr1 and Nr4a2 genes are robustly and specifically expressed in the MHb expressing both the choline acetyltransferase in the ventral part and the neuropeptide SP in the dorsal part [191, 193-195]. In addition, Nurr1 has been reported to mediate a gene pathway involved in habenula development regulated by the POU-domain transcription factor Brn3a [195]. Notably, several studies have suggested that habenular activity is involved in depression, which is one of the most common psychiatric symptoms in AD and a risk factor for AD development. ...
... ]. In addition, Nurr1 has been reported to mediate a gene pathway involved in habenula development regulated by the POU-domain transcription factor Brn3a [195]. Notably, several studies have suggested that habenular activity is involved in depression, which is one of the most common psychiatric symptoms in AD and a risk factor for AD development. ...
Cerebral correlates of depressed behavior in rats, visualized using 14C-2-deoxyglucose autoradiography
1
1988
... In the genetic helpless model and α-methyl-para-tyrosine-induced depression model, brain metabolism and glucose metabolism were elevated in the habenular compared to the control while metabolism of other brain regions was reduced [196, 197]. In patients with depression, it is shown that habenula activity is strongly correlated with dorsal raphé nuclei activity providing an evidence for the important roles of feedback pathway between habenula and dorsal raphé nuclei in controlling release of serotonin [198]. Application of deep brain stimulation to the LHb in a therapy-refractory patient with depression caused successful remission of depression, clarifying the relationship between the habenula and depression [199]. These reports suggest that upregulation of Nurr1 may have a positive effect on not only AD-related pathology but also the psychiatric symptoms that may occur in patients with AD. ...
Opposite metabolic changes in the habenula and ventral tegmental area of a genetic model of helpless behavior
1
2003
... In the genetic helpless model and α-methyl-para-tyrosine-induced depression model, brain metabolism and glucose metabolism were elevated in the habenular compared to the control while metabolism of other brain regions was reduced [196, 197]. In patients with depression, it is shown that habenula activity is strongly correlated with dorsal raphé nuclei activity providing an evidence for the important roles of feedback pathway between habenula and dorsal raphé nuclei in controlling release of serotonin [198]. Application of deep brain stimulation to the LHb in a therapy-refractory patient with depression caused successful remission of depression, clarifying the relationship between the habenula and depression [199]. These reports suggest that upregulation of Nurr1 may have a positive effect on not only AD-related pathology but also the psychiatric symptoms that may occur in patients with AD. ...
Covariation of activity in habenula and dorsal raphe nuclei following tryptophan depletion
1
1999
... In the genetic helpless model and α-methyl-para-tyrosine-induced depression model, brain metabolism and glucose metabolism were elevated in the habenular compared to the control while metabolism of other brain regions was reduced [196, 197]. In patients with depression, it is shown that habenula activity is strongly correlated with dorsal raphé nuclei activity providing an evidence for the important roles of feedback pathway between habenula and dorsal raphé nuclei in controlling release of serotonin [198]. Application of deep brain stimulation to the LHb in a therapy-refractory patient with depression caused successful remission of depression, clarifying the relationship between the habenula and depression [199]. These reports suggest that upregulation of Nurr1 may have a positive effect on not only AD-related pathology but also the psychiatric symptoms that may occur in patients with AD. ...
Remission of major depression under deep brain stimulation of the lateral habenula in a therapy-refractory patient
1
2010
... In the genetic helpless model and α-methyl-para-tyrosine-induced depression model, brain metabolism and glucose metabolism were elevated in the habenular compared to the control while metabolism of other brain regions was reduced [196, 197]. In patients with depression, it is shown that habenula activity is strongly correlated with dorsal raphé nuclei activity providing an evidence for the important roles of feedback pathway between habenula and dorsal raphé nuclei in controlling release of serotonin [198]. Application of deep brain stimulation to the LHb in a therapy-refractory patient with depression caused successful remission of depression, clarifying the relationship between the habenula and depression [199]. These reports suggest that upregulation of Nurr1 may have a positive effect on not only AD-related pathology but also the psychiatric symptoms that may occur in patients with AD. ...
Increased NF-kappaB signalling up-regulates BACE1 expression and its therapeutic potential in Alzheimer's disease
1
2012
... As described so far, Nurr1 has the potential to mitigate the various pathophysiological consequences caused by AD. Nurr1 inhibits NF-κB signaling by binding to and clearance of NF-κB-p65 [79]. Therefore, it can be speculated that modulation of Nurr1 expression can suppress neuroinflammatory responses as well as beta-secretase 1 (BACE1), which is mediated by NF-κB signaling [200]. In addition, Nurr1 plays a neuroprotective role against neuronal death induced by various toxic mediators such as ROS and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [59, 60]. Furthermore, upregulation of Nurr1 has been reported to enhance cognitive function as well as increase hippocampal neurogenesis by enhancing the proliferation and differentiation of NPCs [18]. Considering the beneficial effects of Nurr1 enhancement on the pathological symptoms associated with AD, such as neuroinflammation, neuronal loss, impaired neurogenesis, and cognitive dysfunction, compensation and enhancement of the degenerated Nurr1 in AD patients may be a promising therapeutic target. Remarkably, one recent study reported that Nurr1 regulates AD-related pathogenesis and cognitive function in Aβ-overexpressing mice, supporting the therapeutic potential of Nurr1 for AD [16]. ...
Heterodimerization between members of the Nur subfamily of orphan nuclear receptors as a novel mechanism for gene activation
1
1999
... Interestingly, Nurr1 not only forms homo- or heterodimers with other members of the NR4A family, but can also forms heterodimers with RXRs via DR5 response element [201]. In addition, since the activation of nuclear receptors such as RXR, liver X receptor (LXR), and peroxisome proliferator-activated receptor (PPAR)-γ is known to affect Aβ generation and Aβ clearance, Nurr1 may contribute to the alleviation of Aβ-related pathophysiology through interactions with other nuclear receptors [202-205]. ...
ApoE-directed therapeutics rapidly clear beta-amyloid and reverse deficits in AD mouse models
1
2012
... Interestingly, Nurr1 not only forms homo- or heterodimers with other members of the NR4A family, but can also forms heterodimers with RXRs via DR5 response element [201]. In addition, since the activation of nuclear receptors such as RXR, liver X receptor (LXR), and peroxisome proliferator-activated receptor (PPAR)-γ is known to affect Aβ generation and Aβ clearance, Nurr1 may contribute to the alleviation of Aβ-related pathophysiology through interactions with other nuclear receptors [202-205]. ...
Impact and Therapeutic Potential of PPARs in Alzheimer's Disease
1
2011
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
Mechanisms underlying the rapid peroxisome proliferator-activated receptor-gamma-mediated amyloid clearance and reversal of cognitive deficits in a murine model of Alzheimer's disease
0
2012
Nuclear receptors as therapeutic targets for Alzheimer's disease
2
2011
... Overview of the possible roles of Nurr1 in AD.
References
Nurr1 expression in AD
• Nurr1 immunofluorescence intensity is reduced in the substantia nigra of AD patients
[13]
• Nurr1 mRNA levels are reduced in APPswe, lnd mutant mice
[62, 63]
• The number of Nurr1(+) cells is age-dependently reduced in the subiculum of 5XFAD mice
[90]
• Nurr1 protein is co-localization with Aβ at the early stage in 5XFAD mice
[90]
• Nurr1 protein and mRNA are downregulated in Aβ1-42 fibril-treated CGNs and the hMSC cell line
[65]
Neuroprotective effects
• MPTP-induced neurotoxic vulnerability of dopaminergic neurons is increased in Nurr1(+/-) mice
[59]
• Nurr1 in microglia and astrocytes protects neurons by regulating the production of toxic mediators
[79]
• Ligand and agonist of Nurr1 shows neuroprotective effect against oxidative insult such as MPTP and 6-OHAD
[17, 19, 20]
• Increased expression of Nurr1 upregulates genes involved in ROS detoxification such as Sesn3, Alb2, and Sod1
[81]
• In NSCs, the overexpression of Nurr1 protects against oxidative stress by downregulating cell death-related proteins such as caspase-3 and caspase-11
[60]
• Exogenous Nurr1 induces the differentiation of dopaminergic neurons, and sustained Nurr1 expression improves survival of dopaminergic neurons
[83, 149]
Anti-inflammatory effects
• Nurr1 phosphorylation promotes binding to p65 and recruits the CoREST complex to promoters of inflammatory genes, resulting in inhibition of neuroinflammation
[79]
• Overexpression of Nurr1 suppresses inflammation, whereas knockdown of Nurr1 enhances inflammation
[16]
• NR4A receptors are involved in a negative feedback loop as modulators of the inflammation mechanism
... Interestingly, Nurr1 not only forms homo- or heterodimers with other members of the NR4A family, but can also forms heterodimers with RXRs via DR5 response element [201]. In addition, since the activation of nuclear receptors such as RXR, liver X receptor (LXR), and peroxisome proliferator-activated receptor (PPAR)-γ is known to affect Aβ generation and Aβ clearance, Nurr1 may contribute to the alleviation of Aβ-related pathophysiology through interactions with other nuclear receptors [202-205]. ...
Identification of a potent agonist of the orphan nuclear receptor Nurr1
1
2006
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
Identification of a series of highly potent activators of the Nurr1 signaling pathway
0
2007
Discovery of ligands for Nurr1 by combined use of NMR screening with different isotopic and spin-labeling strategies
1
2007
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
4-amino-7-chloroquinoline derivatives for treating Parkinson's disease: implications for drug discovery
1
2016
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
Effects of isoxazolo-pyridinone 7e, a potent activator of the Nurr1 signaling pathway, on experimental autoimmune encephalomyelitis in mice
0
2014
Development of a novel NURR1/NOT agonist from hit to lead and candidate for the potential treatment of Parkinson's disease
0
2019
Anti-parkinsonian effects of Nurr1 activator in ubiquitin-proteasome system impairment induced animal model of Parkinson's disease
0
2012
Identification of the antineoplastic agent 6-mercaptopurine as an activator of the orphan nuclear hormone receptor Nurr1
1
2003
... Therefore, many researchers have applied various methods to identify potential activators, ligands, and agonists of Nurr1 and have suggested several candidate structures [206-208]. To date, several Nurr1 agonists/activators have been identified and have shown positive effects in autoimmune disease and various PD models, suggesting the potential for the therapeutic effect of Nurr1 in neurodegenerative disease including AD [19, 20, 209-213]. Therefore, a breakthrough for AD treatment will be to prove the efficacy of Nurr1 agonists/mimetics or gene delivery of Nurr1 in animal experiments or clinical trials. A number of studies have suggested that modulation of Nurr1 function may serve as a great strategy to control AD pathogenesis (Fig. 1), and one recent study has shown a colocalization and correlation between Nurr1 and Aβ, and demonstrated that administration of Nurr1 agonists alleviates AD-related pathologies in Aβ-overexpressing mice [16]. ...
Nurr1 is required for maintenance of maturing and adult midbrain dopamine neurons
1
2009
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Dopamine biosynthesis is selectively abolished in substantia nigra/ventral tegmental area but not in hypothalamic neurons in mice with targeted disruption of the Nurr1 gene
1
1998
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Nuclear receptors in neurodegenerative diseases
1
2014
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Parkinson's disease: mechanisms and models
1
2003
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Mutations in NR4A2 associated with familial Parkinson disease
1
2003
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Decreased NURR1 and PITX3 gene expression in Chinese patients with Parkinson's disease
0
2012
Altered NR4A Subfamily Gene Expression Level in Peripheral Blood of Parkinson's and Alzheimer's Disease Patients
1
2016
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Nurr1:RXRalpha heterodimer activation as monotherapy for Parkinson's disease
1
2017
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
Nurr1-Based Therapies for Parkinson's Disease
1
2016
... Nurr1 is known to be a key regulator of the development and maintenance of dopaminergic neurons in the midbrain [7, 78, 214, 215] and plays an important role in inhibiting neuronal death through suppression of inflammatory gene expression in microglia and astrocytes [79, 216]. Therefore, numerous studies have investigated whether Nurr1 may be associated with the pathogenesis of PD, which results from the degeneration of midbrain dopaminergic neurons [217]. In addition to decreased expression of Nurr1 in postmortem brain tissue and the peripheral blood of PD patients, a functional mutation of Nurr1 was found in PD [13, 218-220]. Interestingly, activation of Nurr1 or the Nurr1:RXRα heterodimer by agonists such as AQ, SA00025, and BRF110 has been reported to diminish neuronal loss, neuroinflammation, and behavioral symptoms that occur in 6-hydroxydopamine or MPTP-induced PD models [19, 20, 221, 222]. Remarkably, it has been reported that Nurr1 expression is down-regulated in dopaminergic neurons with NFTs of the SN of AD patients. This results indicate that dysregulation of Nurr1 is associated with tauopathies in the dopaminergic neurons of AD patients [13]. In contrast, a recent study demonstrated that Nurr1 expression is not altered in the SN of postmortem brains of AD patients [16]. This inconsistency may be due to the presence of NFT in the neurons of AD brains. As Nurr1 expression levels were not altered in neurons without NFT [13], although the latter study did not provide further information regarding the presence of NFT in the neurons [16]. These data provide evidence that Nurr1 may be a relevant target for alleviating AD pathogenesis, especially in tauopathy. ...
miR-145-5p/Nurr1/TNF-alpha Signaling-Induced Microglia Activation Regulates Neuron Injury of Acute Cerebral Ischemic/Reperfusion in Rats
1
2017
... A recent study has shown that Nurr1 expression was dynamic following acute ischemia induced by middle cerebral artery occlusion (MCAO)/reperfusion in a rat model. There was a negative correlation between Nurr1 and infarct volume up to 12 hours after ligation, but a positive correlation was observed after 24 hours. Nurr1 overexpression inhibited tumor necrosis factor-α (TNF-α) levels in microglia. Increase of Nurr1 expression through suppression of miR-145-5p, a negative regulator of Nurr1 alleviated infarct volume and improved the neurological outcomes in an acute stroke model [223]. In addition, transplantation of Nurr1-overexpressing human embryonal carcinoma cells into the ischemic striatum restored the behavioral disorder in a transient MCAO rat model [153]. Considering that TNF-α released from Aβ-activated microglia is a key cytokine causing cell cycle events, which are related to pathogenesis of neuronal death in AD [224], these regulatory effects of Nurr1 on microglial activation could be a therapeutic target for AD as well as ischemic stroke. ...
... A recent study has shown that Nurr1 expression was dynamic following acute ischemia induced by middle cerebral artery occlusion (MCAO)/reperfusion in a rat model. There was a negative correlation between Nurr1 and infarct volume up to 12 hours after ligation, but a positive correlation was observed after 24 hours. Nurr1 overexpression inhibited tumor necrosis factor-α (TNF-α) levels in microglia. Increase of Nurr1 expression through suppression of miR-145-5p, a negative regulator of Nurr1 alleviated infarct volume and improved the neurological outcomes in an acute stroke model [223]. In addition, transplantation of Nurr1-overexpressing human embryonal carcinoma cells into the ischemic striatum restored the behavioral disorder in a transient MCAO rat model [153]. Considering that TNF-α released from Aβ-activated microglia is a key cytokine causing cell cycle events, which are related to pathogenesis of neuronal death in AD [224], these regulatory effects of Nurr1 on microglial activation could be a therapeutic target for AD as well as ischemic stroke. ...
Reduction of dopamine-related transcription factors Nurr1 and NGFI-B in the prefrontal cortex in schizophrenia and bipolar disorders
1
2006
... Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
Pathway-Specific Dopamine Abnormalities in Schizophrenia
2
2017
... Nurr1 heterozygous mice exhibited behavioral patterns associated with the symptoms of schizophrenia and were suggested as a potential animal model of schizophrenia [15, 51]. Furthermore, protein and mRNA expression levels of Nurr1 were reduced in the prefrontal cortex of schizophrenia patients [225]. Considering that the abnormal function of dopaminergic neurons in the cerebral cortex and subcortical areas is associated with schizophrenia [226], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
... ], the changes in Nurr1 expression in schizophrenia address necessitate studies examining the correlation between schizophrenia and Nurr1. Despite that hyperactivation of dopaminergic neurotransmission, conventionally considered a major hypothesis for pathology schizophrenia, these findings may support the recent challenges against the conventional dopamine hypothesis [226]. In relation to changes in Nurr1 expression in AD, the reduced number of Nurr1-expressing cells in subiculum of AD model mice with disease progress [64] and decreased levels of dopamine in various regions including the hippocampus of Nurr1 heterozygous mice, a schizophrenic animal model [15], may demonstrate some similarities in molecular changes involving both disorders as well as provide additional insights for further studies comparing mechanisms between two diseases. ...
Opposite effects of acute and chronic amphetamine on Nurr1 and NF-kappaB p65 in the rat ventral tegmental area
1
2016
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Chronic cocaine administration modulates the expression of transcription factors involved in midbrain dopaminergic neuron function
0
2007
Heroin abuse is characterized by discrete mesolimbic dopamine and opioid abnormalities and exaggerated nuclear receptor-related 1 transcriptional decline with age
0
2007
Chronic methamphetamine administration causes differential regulation of transcription factors in the rat midbrain
1
2011
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Decreased level of Nurr1 in heterozygous young adult mice leads to exacerbated acute and long-term toxicity after repeated methamphetamine exposure
1
2010
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Rhynchophylline Downregulates Phosphorylated cAMP Response Element Binding Protein, Nuclear Receptor-related-1, and Brain-derived Neurotrophic Factor Expression in the Hippocampus of Ketamine-induced Conditioned Place Preference Rats
1
2018
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Effects of rhynchophylline on the hippocampal miRNA expression profile in ketamine-addicted rats
1
2018
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
Reward processing in neurodegenerative disease
1
2015
... Although the role of Nurr1 in addiction is controversial, depending on the duration of treatment and the drug used, a number of studies have reported that the administration of addictive drugs such as cocaine and heroin reduces Nurr1 transcript levels in the midbrain [227-230]. In addition, Nurr1 heterozygous mice exhibited reduced reward-seeking behaviors mediated by dopaminergic neurotransmission and were vulnerable to neuro-degeneration during long-term methamphetamine administration [14, 231]. In contrast, the hippocampus of ketamine-addicted rats has been reported to show increased levels of Nurr1 due to CREB-medicated phosphorylation [232, 233]. These studies suggest that Nurr1 is involved in the initiation and progression of addictive disorders, which may depend on the type of drug and the duration of administration. As several lines of evidence suggest that reward processing is defective in neurodegenerative diseases including AD [234], the correlation between Nurr1 expression and addictive behaviors may suggest the need for further investigation regarding the relationship between Nurr1 function in AD and addictive behaviors. ...
A selective group of dopaminergic neurons express Nurr1 in the adult mouse brain
1
1999
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Immunocytochemical expression of dopamine-related transcription factors Pitx3 and Nurr1 in prenatally stressed adult rats
1
2009
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
The Role of Dopamine and Its Dysfunction as a Consequence of Oxidative Stress
1
2016
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Joint analysis of the DRD5 marker concludes association with attention-deficit/hyperactivity disorder confined to the predominantly inattentive and combined subtypes
1
2004
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Dopamine system genes and attention deficit hyperactivity disorder: a meta-analysis
1
2002
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Evaluating dopamine reward pathway in ADHD: clinical implications
1
2009
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Prenatal immune activation interacts with genetic Nurr1 deficiency in the development of attentional impairments
1
2012
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
The dual roles of cytokines in Alzheimer's disease: update on interleukins, TNF-alpha, TGF-beta and IFN-gamma
1
2016
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Identification and characterization of human NR4A2 polymorphisms in attention deficit hyperactivity disorder
1
2005
... VTA, a dopamine nucleus brain region with robust expression of Nurr1 [235, 236], projects dopamine axons to the prefrontal cortex [237], and dopamine system is considered to be an important part of ADHD pathogenesis [238, 239]. In addition, decreased dopamine synaptic markers have been reported in the dopamine reward pathway in ADHD patients [240]. An in vivo study using Nurr1 knockout mice with prenatal immune activation as an attention impairment model reported that genetic and environmental factors synergistically affected attentional impairment as well as additively affected locomotor hyperactivity. Remarkably, Nurr1 heterodeficient mice showed increased locomotor activity, and exhibited altered inflammatory cytokine responses against prenatal immune activation [241]. Although there were inconsistent reports regarding the levels of cytokines in AD, cytokines such as IL-6 and IL-10 are known to play important roles in AD pathogenesis [242]. Given that two NR4A2 polymorphisms were found in patients with ADHD [243], further studies investigating the changes of Nurr1 expression in ADHD may provide a better understanding of AD pathogenesis. ...
Sleep and circadian rhythm disruption in psychiatric and neurodegenerative disease
1
2010
... Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD. ...
Sleep and circadian rhythm disruption and stress intersect in Alzheimer's disease
1
2019
... Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD. ...
Sleep, circadian rhythms, and the pathogenesis of Alzheimer disease
1
2015
... Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD. ...
Impact of circadian nuclear receptor REV-ERBalpha on midbrain dopamine production and mood regulation
1
2014
... Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD. ...
Alterations of Clock Gene RNA Expression in Brain Regions of a Triple Transgenic Model of Alzheimer's Disease
1
2017
... Disorders of the midbrain dopaminergic neurons, which are the basis of the reward system in the brain, are involved in the disruption of the circadian rhythm [244]. Notably, sleep and circadian rhythm disorder are early biomarkers of AD [245, 246]. The circadian nuclear receptor REV-ERBα encoded by the NR1D1 gene, competes with Nurr1 for the regulation of circadian TH expression via a target-dependent antagonistic mechanism [247]. In 6-month old 3xTg-AD mice, an animal model of AD, gene expression of NR1D1 is increased in the brainstem after exposure to darkness, compared to control mice [248]. Thus, these data may imply that controlling the balance between expression of ERV-ERBα and Nurr1 could be a potential target for treating circadian rhythm disorder in AD. ...
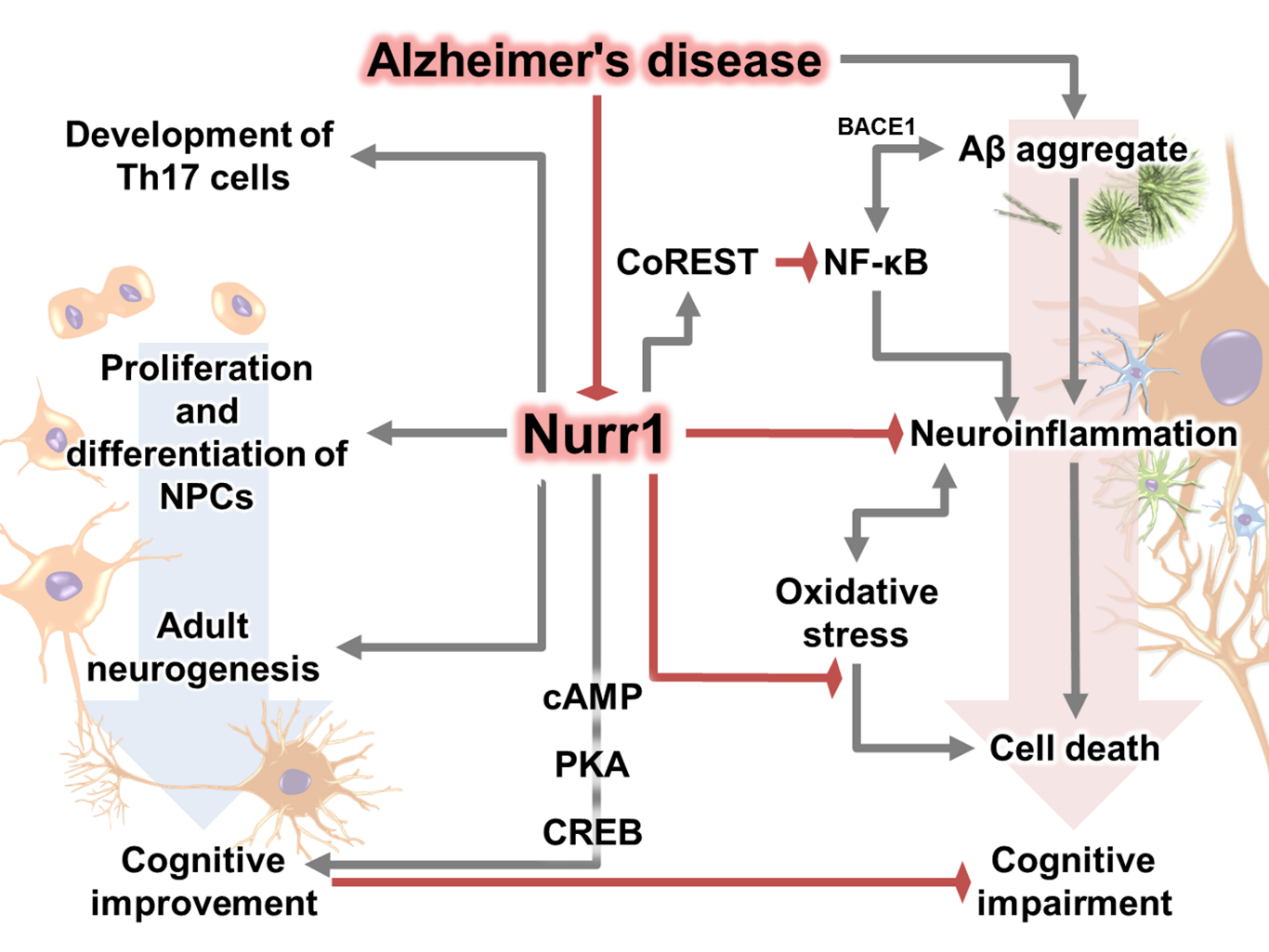
{kind=link}
{kind=link}



