1Institute of Biology of FRC of Komi Scientific Center of Ural Branch of Russian Academy of Sciences, Syktyvkar, 167982, Russia.2Vetek (Seniority), The Movement for Longevity and Quality of Life, Israel.3Laboratory of Immunopathology and Immunosenescence, Department of Biomedicine, Neurosciences and Advanced Diagnostics, University of Palermo, Palermo, Italy
通讯作者: Correspondence should be addressed to: Dr. Alexey Moskalev, Institute of Biology of FRC of Komi Scientific Center of Ural Branch of Russian Academy of Sciences, Russia. Email: amoskalev@list.ru; Dr. Ilia Stambler, The Movement for Longevity and Quality of Life, Israel. Email: ilia.stambler@gmail.com; Dr. Calogero Caruso, University of Palermo, Palermo, Italy. Email: calogero.caruso@unipa.it.Correspondence should be addressed to: Dr. Alexey Moskalev, Institute of Biology of FRC of Komi Scientific Center of Ural Branch of Russian Academy of Sciences, Russia. Email: amoskalev@list.ru; Dr. Ilia Stambler, The Movement for Longevity and Quality of Life, Israel. Email: ilia.stambler@gmail.com; Dr. Calogero Caruso, University of Palermo, Palermo, Italy. Email: calogero.caruso@unipa.it.Correspondence should be addressed to: Dr. Alexey Moskalev, Institute of Biology of FRC of Komi Scientific Center of Ural Branch of Russian Academy of Sciences, Russia. Email: amoskalev@list.ru; Dr. Ilia Stambler, The Movement for Longevity and Quality of Life, Israel. Email: ilia.stambler@gmail.com; Dr. Calogero Caruso, University of Palermo, Palermo, Italy. Email: calogero.caruso@unipa.it.Correspondence should be addressed to: Dr. Alexey Moskalev, Institute of Biology of FRC of Komi Scientific Center of Ural Branch of Russian Academy of Sciences, Russia. Email: amoskalev@list.ru; Dr. Ilia Stambler, The Movement for Longevity and Quality of Life, Israel. Email: ilia.stambler@gmail.com; Dr. Calogero Caruso, University of Palermo, Palermo, Italy. Email: calogero.caruso@unipa.it.
收稿日期:2020-05-4
修回日期: 2020-06-1
接受日期: 2020-06-3
网络出版日期: 2020-12-01
版权声明:
2020 this is an open access article distributed under the terms of the creative commons attribution license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
展开
Abstract
The interrelation of the processes of immunity and senescence now receives an unprecedented emphasis during the COVID-19 pandemic, which brings to the fore the critical need to combat immunosenescence and improve the immune function and resilience of older persons. Here we review the historical origins and the current state of the science of innate and adaptive immunity in aging and longevity. From the modern point of view, innate and adaptive immunity are not only affected by aging but also are important parts of its underlying mechanisms. Excessive levels or activity of antimicrobial peptides, C-reactive protein, complement system, TLR/NF-κB, cGAS/STING/IFN 1,3 and AGEs/RAGE pathways, myeloid cells and NLRP3 inflammasome, declined levels of NK cells in innate immunity, thymus involution and decreased amount of naive T-cells in adaptive immunity, are biomarkers of aging and predisposition factors for cellular senescence and aging-related pathologies. Long-living species, human centenarians, and women are characterized by less inflamm-aging and decelerated immunosenescence. Despite recent progress in understanding, the harmonious theory of immunosenescence is still developing. Geroprotectors targeting these mechanisms are just emerging and are comprehensively discussed in this article.
MoskalevAlexey, StamblerIlia, CarusoCalogero. Innate and Adaptive Immunity in Aging and Longevity: The Foundation of Resilience[J]. Aging and Disease, 2020, 11(6): 1363-1373 https://doi.org/10.14336/AD.2020.0603
The interrelation of the processes of immunity and senescence now receives an unprecedented emphasis during the COVID-19 pandemic, which has stressed the critical need to combat immunosenescence and improve the immune function and resilience of older persons. At this time, it is appropriate to review the current state of the science of innate and adaptive immunity in aging and longevity, as well as the historical origins of this field of study, to further promote the research in this area. That is the subject of the present work.
Historically, the field originated at the turn of the 20th century with the work of Elie Metchnikoff whose 175th anniversary we celebrated on May 15, 2020 (May 15, 1845 - July 15, 1916). Metchnikoff is well recognized as a pioneering immunologist and microbiologist, a vice-director of the Pasteur Institute in Paris, and the Nobel Laureate in Physiology or Medicine of 1908 for the discovery of phagocytosis (a major contribution to the cellular theory of immunity). Yet, he may also be well credited as “the father” of gerontology - the disciplinary term he coined. Both the terms “gerontology” (“the study of aging”) and “thanatology” (“the study of death”) were coined by him in the “Etudes On the Nature of Man” published in 1903, which may mark the beginning of these scientific fields. Moreover, Metchnikoff can also be credited for the establishment of the interdisciplinary connection between these fields, in particular between aging research and immunology. Metchnikoff was the author of arguably the first systematic scientific theory of aging, interrelating the processes of immunity and senescence (www.longevityhistory.com/) [1]. In Metchnikoff’s own words: “We saw that, during aging, there occurs a struggle between noble elements (parenchymal tissues, e.g. the tissues of the muscle, kidney, lung, and brain) and phagocytes (“low/primitive elements”), and that the vitality of the former is, for the most part, diminished, whereas the latter, on the contrary, show increased activity. Therefore, it would seem that the means to use in the struggle against pathological aging should be, on the one hand, the strengthening of the most valuable elements of the organism, and on the other, the attenuation of the aggressive onslaught of the phagocytes. I must point out to the reader from the beginning that this problem is not yet solved, but its solution does not involve anything impossible. It is a scientific question, like many others.” (Elie Metchnikoff, Etudy o Prirode Cheloveka (Etudes on the Nature of Man), The USSR Academy of Sciences Press, Moscow, 1961 (1903), Ch. X. “Vvedenie v nauchnoe izuchenie starosti” (An introduction to the scientific study of aging), pp. 201-202.)
Following a century of study, at the present time, natural immunity is understood to consist of three interrelated parts: physiological barriers, innate immunity and adaptive immunity. All of these are affected by aging [2]. Immunosenescence results in increased susceptibility and severity of infectious diseases and non-communicable age-associated diseases, among them cancer, cardio-vascular disease, and autoimmunity [3].
The molecular mechanisms of the induction of inflammation and cellular senescence intersect through activation of the TLR/NF-κB, cGAS/STING/IFN 1,3, AGEs/RAGE molecular signaling pathways and the assembly of the NLRP3 inflammasome. Chronic sterile inflammation with aging was termed by Claudio Franceschi “inflamm-aging” [4]. The hyperactivation of the innate immunity response predominantly reduces the lifespan.
Antimicrobial peptides are involved in chemotaxis and activation of innate and adaptive immunity cells in different animals from invertebrates to humans [5, 6]. On the model of Drosophila, it is shown that hyperactivation of different antimicrobial peptide genes significantly reduces the lifespan [7].
Pathogen-associated molecular patterns of microorganisms are recognized by the innate immune system through inherited pattern-recognition receptors [8], including C-reactive protein [9], Toll-like-receptors [10] and some cytoplasmic receptors, including cGas/STING [11].
Figure 1. Gut dysbiosis/permeability with aging can induce TLR in microglia and exerts mitochondrial dysfunction.
C-reactive protein (CRP) is a soluble pathogen pattern recognition receptor. It binds to 1,6-Bis(phosphocholine) of cell membranes of damaged cells or bacteria to induce complement or immune cell activation [9]. CRP hyperactivity is a biomarker of aging and is connected with inflammation and fibrosis [12].
The complement system includes more than 50 proteins in the plasma and cell membrane that act in response to activation of pattern recognition receptors, including CRP, killing microbes, sending danger signals, and accelerating apoptosis of damaged cells. There was established the participation of the complement system in the pathogenesis of aging-dependent diseases and their complications, including age-related macular degeneration [13] and type 2 diabetes [14].
Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19].
Figure 2. AGEs/RAGE pathway and age-related diseases.
cGas/STING pathway is the main intracellular sensor of viral invasion, including SARS-Cov-2, to induce Interferon 1 and 3 productions [20]. The STING pathway is also hyperactivated with aging by internal reasons, including retrotransposons, chromatin and mtDNA fragments in the cytosol with a consequent interferon induction, cellular senescence and apoptosis [21-24]. This could be a hypothetical reason for the greater severity of Covid-19 in elderly people.
Some simple sugars (glucose, fructose) in the presence of transition metal ions (iron and copper) react chemically with amino acid residues (lysine, arginine) in proteins, such as collagen and elastin, causing the formation of glycation end products that not only increase the extracellular matrix stiffness, but also induce chronic inflammation through their RAGE receptors on the surface of cells (Fig. 2), such as vascular endothelium [25].
Cellular senescence itself can induce inflammation by secreting pro-inflammatory cytokines, the so-called Senescence Associated Secretory Phenotype (SASP) [26] (Fig. 3).
According to Baker's study, elimination of senescent cells prolongs the lifespan and healthspan of mice [27, 28].
Figure 3. SASP involvement in aging-related pathologies.
Adiposity is another source of inflamm-aging. Adipocyte hypertrophy leads to the secretion of proinflammatory leptin, lipocalin-2, progranulin, and chemoattractants for T- and B-cells [29-33] (Fig. 4).
In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5).
In our experiments on the Drosophila model, we inhibited the different underlying pathways of NF-kB. In most cases, this led to an increase in lifespan [43-45].
It is worth noting that many substances of natural origin contained in food can inhibit NF-kB [46].
Aging is accompanied by gut microbiota alteration, like decreased overall diversity and an increased abundance of proinflammatory species, that can be a part of systemic inflammation and many aging-related diseases [47, 48].
Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55].
Figure 5. Physiological stress factors can induce NF-kB pathway.
Immune parameters associated with survival may vary in diverse populations of different ages. Therefore, we have to focus on the changes considered to be the hallmarks of immunosenescence, based on the literature data. The hallmarks of immunosenescence include: (i) a reduced ability to respond to new antigens; (ii) the accumulation of memory T cells; (iii) a lingering level of low-grade inflammation termed “inflamm-aging.” Mechanistically, immunosenescence is only partially explained by organismal and cellular senescence. Therefore, these hallmarks of immunosenescence would be markedly affected by the history of individual exposure to pathogens. In fact, several factors, such as genetics, nutrition, exercise, previous exposure to microorganisms, biological and cultural sex, and human cytomegalovirus (HCMV) status can influence immunosenescence [56].
Concerning sex/gender, in Western countries, women live 5-6 years more than men do. Furthermore, 85% of over 100 years old are women. It is debated whether women live longer than men for reasons of gender or sex, e.g., for cultural or biological differences. However, females live longer than males in other animal species. There is sexual dimorphism in the immune response, i.e. females are more resistant to infections, but they have a higher incidence of autoimmune diseases compared to males, yet their relevance for life span is negligible. However, age-related changes in various immunological parameters differ between men and women. Findings indicate that the slower rate of decline in immunological parameters in women than those in men is consistent with the fact that women live longer, than men do [57].
Concerning HCMV, virus status has a greater impact than age on the immune system because the virus contributes to shaping the immune profile and function during normal human aging. HCMV seropositivity is closely related to the reversal of the CD4/CD 8 T-cell ratio. In fact, persistent HCMV infection leads to chronic stimulation of CD8 T cells, which expand clonally showing an effector memory phenotype characterized by low CD28 expression. The absolute increase in memory T cells, called memory inflation, is observed only in older people infected by HCMV [58].
One of the pillars of adaptive immunity is the thymus. After an active period of creation and training of new T cells in childhood, at the time of puberty, the thymus undergoes involution, losing the stromal part and filling with fat [59], that can decrease T-cell repertoire to new antigens, including SARS-CoV-2. The involution is continuing during aging, because Wnt4 expression is down-regulated, while their Frizzled receptors and PPARgamma expression increases in the thymus [60]. On the contrary, peripheral T-cell numbers are maintained through the antigen-independent homeostatic proliferation of naive T cells that may lead to the emergence of dysfunctional memory-phenotype CD4+ T cell subpopulation (cell senescence-associated T cells, SA-T cells) [61]. SA-T cells secrete abundant pro-inflammatory factors such as osteopontin and chemokines, playing a direct role in SASP [62].
CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6).
Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74].
T follicular helper (TFH) cells are presented in lymphoid organs and in peripheral blood and help B cells for the production of immunoglobulins. Dysfunctional TFH cells with aging play a role in cancer, autoimmune and cardiovascular diseases [75].
Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80].
Human centenarians are a model for healthy aging. The longest living cohort of Italian centenarians has more favorable values of important immune parameters: naïve, activated/memory and effector/memory T cells [81]. Healthy centenarians presented with a distinct expression of proteins/pathways that reflect a healthy immune function, including less inflamm-aging and autoimmunity and increased B cell-mediated immune response [82].
Figure 7. Changes occurring during aging (reproduced with permission from [56]).
Centenarians have been used as an optimal model for successful aging. However, this model shows several limitations, in particular the selection of appropriate controls. Thus, the interest has been centered on centenarian offspring, since it is well known that they are healthier than the remaining old people are. Accordingly, significant differences between old subjects and centenarian offspring, in most of the studied T and B subsets, show that centenarian offspring subsets present intermediate phenotyping between old and younger people. Therefore, centenarian offspring retain more youthful immunological parameters and the exhaustion of the immune system is less evident than in old people without centenarian parents [83]. Therefore, cell subset changes could represent a hallmark of successful or unsuccessful aging and could be used as a biomarker of human life span, potentially useful for the evaluation of immunosenescence treatment [83, 84].
Despite recent progress in understanding (Fig. 7), the harmonious theory of immunosenescence is still developing.
Based on the present level of knowledge, the geroprotective therapies targeting the mechanisms of immunosenescence are just emerging [85]. Their studies need to intensify, with a broader identification of potential clinically applicable interventions and biomarkers, and their extensive pre-clinical and clinical testing [85].
There are several hundreds of potential geroprotective interventions, that have been demonstrated on model organisms and collected in online databases Geroprotectors.Org [86] and DrugAge [87]. Not all of them meet the criteria of effective and safe treatment applicable for humans [88, 89].
According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94].
Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99].
Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation.
Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115].
Gaining this knowledge is urgently needed to enhance the quality of life and health span of the global aging population, to improve their resilience against both non-communicable and communicable diseases.
Multiple Sclerosis: Melatonin, Orexin, and Ceramide Interact with Platelet Activation Coagulation Factors and Gut-Microbiome-Derived Butyrate in the Circadian Dysregulation of Mitochondria in Glia and Immune Cells
Regulation of Toll-Like Receptor (TLR) Signaling Pathway by Polyphenols in the Treatment of Age-Linked Neurodegenerative Diseases: Focus on TLR4 Signaling
Superiority of sirolimus (rapamycin) over cyclosporine in augmenting allograft and xenograft survival in mice treated with antilymphocyte serum and donor-specific bone marrow
KraigE, LinehanLA, LiangH, RomoTQ, LiuQ, WuY, et al. (2018).
A randomized control trial to establish the feasibility and safety of rapamycin treatment in an older human cohort: Immunological, physical performance, and cognitive effects
Elie Metchnikoff-The founder of longevity science and a founder of modern medicine: In honor of the 170th anniversary
1
2015
... Historically, the field originated at the turn of the 20th century with the work of Elie Metchnikoff whose 175th anniversary we celebrated on May 15, 2020 (May 15, 1845 - July 15, 1916). Metchnikoff is well recognized as a pioneering immunologist and microbiologist, a vice-director of the Pasteur Institute in Paris, and the Nobel Laureate in Physiology or Medicine of 1908 for the discovery of phagocytosis (a major contribution to the cellular theory of immunity). Yet, he may also be well credited as “the father” of gerontology - the disciplinary term he coined. Both the terms “gerontology” (“the study of aging”) and “thanatology” (“the study of death”) were coined by him in the “Etudes On the Nature of Man” published in 1903, which may mark the beginning of these scientific fields. Moreover, Metchnikoff can also be credited for the establishment of the interdisciplinary connection between these fields, in particular between aging research and immunology. Metchnikoff was the author of arguably the first systematic scientific theory of aging, interrelating the processes of immunity and senescence (www.longevityhistory.com/) [1]. In Metchnikoff’s own words: “We saw that, during aging, there occurs a struggle between noble elements (parenchymal tissues, e.g. the tissues of the muscle, kidney, lung, and brain) and phagocytes (“low/primitive elements”), and that the vitality of the former is, for the most part, diminished, whereas the latter, on the contrary, show increased activity. Therefore, it would seem that the means to use in the struggle against pathological aging should be, on the one hand, the strengthening of the most valuable elements of the organism, and on the other, the attenuation of the aggressive onslaught of the phagocytes. I must point out to the reader from the beginning that this problem is not yet solved, but its solution does not involve anything impossible. It is a scientific question, like many others.” (Elie Metchnikoff, Etudy o Prirode Cheloveka (Etudes on the Nature of Man), The USSR Academy of Sciences Press, Moscow, 1961 (1903), Ch. X. “Vvedenie v nauchnoe izuchenie starosti” (An introduction to the scientific study of aging), pp. 201-202.) ...
The Immune System and Its Dysregulation with Aging
1
2019
... Following a century of study, at the present time, natural immunity is understood to consist of three interrelated parts: physiological barriers, innate immunity and adaptive immunity. All of these are affected by aging [2]. Immunosenescence results in increased susceptibility and severity of infectious diseases and non-communicable age-associated diseases, among them cancer, cardio-vascular disease, and autoimmunity [3]. ...
Does the human immune system ever really become "senescent"?
1
2017
... Following a century of study, at the present time, natural immunity is understood to consist of three interrelated parts: physiological barriers, innate immunity and adaptive immunity. All of these are affected by aging [2]. Immunosenescence results in increased susceptibility and severity of infectious diseases and non-communicable age-associated diseases, among them cancer, cardio-vascular disease, and autoimmunity [3]. ...
Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans
1
2007
... The molecular mechanisms of the induction of inflammation and cellular senescence intersect through activation of the TLR/NF-κB, cGAS/STING/IFN 1,3, AGEs/RAGE molecular signaling pathways and the assembly of the NLRP3 inflammasome. Chronic sterile inflammation with aging was termed by Claudio Franceschi “inflamm-aging” [4]. The hyperactivation of the innate immunity response predominantly reduces the lifespan. ...
Antimicrobial Peptides: An Emerging Category of Therapeutic Agents
1
2016
... Antimicrobial peptides are involved in chemotaxis and activation of innate and adaptive immunity cells in different animals from invertebrates to humans [5, 6]. On the model of Drosophila, it is shown that hyperactivation of different antimicrobial peptide genes significantly reduces the lifespan [7]. ...
Defensins in innate immunity
1
2014
... Antimicrobial peptides are involved in chemotaxis and activation of innate and adaptive immunity cells in different animals from invertebrates to humans [5, 6]. On the model of Drosophila, it is shown that hyperactivation of different antimicrobial peptide genes significantly reduces the lifespan [7]. ...
Overexpression of antimicrobial peptides contributes to aging through cytotoxic effects in Drosophila tissues
1
2018
... Antimicrobial peptides are involved in chemotaxis and activation of innate and adaptive immunity cells in different animals from invertebrates to humans [5, 6]. On the model of Drosophila, it is shown that hyperactivation of different antimicrobial peptide genes significantly reduces the lifespan [7]. ...
Pathogen Recognition and Innate Immunity
1
2006
... Pathogen-associated molecular patterns of microorganisms are recognized by the innate immune system through inherited pattern-recognition receptors [8], including C-reactive protein [9], Toll-like-receptors [10] and some cytoplasmic receptors, including cGas/STING [11]. ...
Targeting C-Reactive Protein in Inflammatory Disease by Preventing Conformational Changes
2
2015
... Pathogen-associated molecular patterns of microorganisms are recognized by the innate immune system through inherited pattern-recognition receptors [8], including C-reactive protein [9], Toll-like-receptors [10] and some cytoplasmic receptors, including cGas/STING [11]. ...
... C-reactive protein (CRP) is a soluble pathogen pattern recognition receptor. It binds to 1,6-Bis(phosphocholine) of cell membranes of damaged cells or bacteria to induce complement or immune cell activation [9]. CRP hyperactivity is a biomarker of aging and is connected with inflammation and fibrosis [12]. ...
Toll-like receptors in immunity and inflammatory diseases: Past, present, and future
1
2018
... Pathogen-associated molecular patterns of microorganisms are recognized by the innate immune system through inherited pattern-recognition receptors [8], including C-reactive protein [9], Toll-like-receptors [10] and some cytoplasmic receptors, including cGas/STING [11]. ...
Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway
1
2013
... Pathogen-associated molecular patterns of microorganisms are recognized by the innate immune system through inherited pattern-recognition receptors [8], including C-reactive protein [9], Toll-like-receptors [10] and some cytoplasmic receptors, including cGas/STING [11]. ...
C-reactive protein and ageing
1
2017
... C-reactive protein (CRP) is a soluble pathogen pattern recognition receptor. It binds to 1,6-Bis(phosphocholine) of cell membranes of damaged cells or bacteria to induce complement or immune cell activation [9]. CRP hyperactivity is a biomarker of aging and is connected with inflammation and fibrosis [12]. ...
Complement system and age-related macular degeneration: drugs and challenges
1
2019
... The complement system includes more than 50 proteins in the plasma and cell membrane that act in response to activation of pattern recognition receptors, including CRP, killing microbes, sending danger signals, and accelerating apoptosis of damaged cells. There was established the participation of the complement system in the pathogenesis of aging-dependent diseases and their complications, including age-related macular degeneration [13] and type 2 diabetes [14]. ...
Complement activation in obesity, insulin resistance, and type 2 diabetes mellitus
1
2020
... The complement system includes more than 50 proteins in the plasma and cell membrane that act in response to activation of pattern recognition receptors, including CRP, killing microbes, sending danger signals, and accelerating apoptosis of damaged cells. There was established the participation of the complement system in the pathogenesis of aging-dependent diseases and their complications, including age-related macular degeneration [13] and type 2 diabetes [14]. ...
Toll-like receptors in the pathogenesis of neuroinflammation
1
2019
... Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
A Dietary Mixture of Oxysterols Induces In Vitro Intestinal Inflammation through TLR2/4 Activation: The Protective Effect of Cocoa Bean Shells
2
2019
... Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
... ). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
New mitochondrial DNA synthesis enables NLRP3 inflammasome activation
1
2018
... Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
Multiple Sclerosis: Melatonin, Orexin, and Ceramide Interact with Platelet Activation Coagulation Factors and Gut-Microbiome-Derived Butyrate in the Circadian Dysregulation of Mitochondria in Glia and Immune Cells
1
2019
... Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
Regulation of Toll-Like Receptor (TLR) Signaling Pathway by Polyphenols in the Treatment of Age-Linked Neurodegenerative Diseases: Focus on TLR4 Signaling
1
2019
... Toll-like receptors (TLR) cell surface receptors can recognize pathogen patterns from viruses, bacteria, or fungi to induce NF-kB proinflammatory signaling. TLR inhibition is a potential target to alleviate neuroinflammation [15]. TLR4 is of particular interest in connection with aging, since it can be activated by cytotoxic oxysterols (7-ketocholesterol), which are formed in tissues during inflammation or come from long-stored food [16]. Its activation leads to mitochondrial dysfunction and an inflammatory reaction [17], including in brain microglia [18] (Fig. 1). TLR4 is inhibited by substances from cocoa [16]. Some polyphenols can suppress overexpression of inflammatory mediators through TLR4/NF-κB/STAT signaling intervention [19]. ...
Immunology of COVID-19: current state of the science
1
2020
... cGas/STING pathway is the main intracellular sensor of viral invasion, including SARS-Cov-2, to induce Interferon 1 and 3 productions [20]. The STING pathway is also hyperactivated with aging by internal reasons, including retrotransposons, chromatin and mtDNA fragments in the cytosol with a consequent interferon induction, cellular senescence and apoptosis [21-24]. This could be a hypothetical reason for the greater severity of Covid-19 in elderly people. ...
Mitochondria-to-nucleus retrograde signaling drives formation of cytoplasmic chromatin and inflammation in senescence
1
2020
... cGas/STING pathway is the main intracellular sensor of viral invasion, including SARS-Cov-2, to induce Interferon 1 and 3 productions [20]. The STING pathway is also hyperactivated with aging by internal reasons, including retrotransposons, chromatin and mtDNA fragments in the cytosol with a consequent interferon induction, cellular senescence and apoptosis [21-24]. This could be a hypothetical reason for the greater severity of Covid-19 in elderly people. ...
LINE1 Derepression in Aged Wild-Type and SIRT6-Deficient Mice Drives Inflammation
0
2019
L1 drives IFN in senescent cells and promotes age-associated inflammation
0
2019
Extranuclear DNA accumulates in aged cells and contributes to senescence and inflammation
1
2019
... cGas/STING pathway is the main intracellular sensor of viral invasion, including SARS-Cov-2, to induce Interferon 1 and 3 productions [20]. The STING pathway is also hyperactivated with aging by internal reasons, including retrotransposons, chromatin and mtDNA fragments in the cytosol with a consequent interferon induction, cellular senescence and apoptosis [21-24]. This could be a hypothetical reason for the greater severity of Covid-19 in elderly people. ...
Role of advanced glycation end products in cellular signaling
1
2014
... Some simple sugars (glucose, fructose) in the presence of transition metal ions (iron and copper) react chemically with amino acid residues (lysine, arginine) in proteins, such as collagen and elastin, causing the formation of glycation end products that not only increase the extracellular matrix stiffness, but also induce chronic inflammation through their RAGE receptors on the surface of cells (Fig. 2), such as vascular endothelium [25]. ...
The senescence-associated secretory phenotype: the dark side of tumor suppression
1
2010
... Cellular senescence itself can induce inflammation by secreting pro-inflammatory cytokines, the so-called Senescence Associated Secretory Phenotype (SASP) [26] (Fig. 3). ...
Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline
1
2018
... According to Baker's study, elimination of senescent cells prolongs the lifespan and healthspan of mice [27, 28]. ...
... According to Baker's study, elimination of senescent cells prolongs the lifespan and healthspan of mice [27, 28]. ...
Adipokines and inflammation: is it a question of weight?
1
2018
... Adiposity is another source of inflamm-aging. Adipocyte hypertrophy leads to the secretion of proinflammatory leptin, lipocalin-2, progranulin, and chemoattractants for T- and B-cells [29-33] (Fig. 4). ...
Obesity decreases B cell responses in young and elderly individuals
0
2016
Obesity and inflammation: the effects of weight loss
0
2008
Adipocytokines: mediators linking adipose tissue, inflammation and immunity
0
2006
Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells
1
2006
... Adiposity is another source of inflamm-aging. Adipocyte hypertrophy leads to the secretion of proinflammatory leptin, lipocalin-2, progranulin, and chemoattractants for T- and B-cells [29-33] (Fig. 4). ...
Inflammatory cause of metabolic syndrome via brain stress and NF-kappaB
1
2012
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Disruption of NF-kappaB signalling by ancient microbial molecules: novel therapies of the future?
1
2010
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Nuclear factor-kappaB is a critical mediator of stress-impaired neurogenesis and depressive behavior
1
2010
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Vitamin D deficiency enhances insulin resistance by promoting inflammation in type 2 diabetes
1
2019
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Core circadian protein CLOCK is a positive regulator of NF-kappaB-mediated transcription
1
2012
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Aldosterone, oxidative stress, and NF-kappaB activation in hypertension-related cardiovascular and renal diseases
1
2012
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Angiotensin II activates nuclear transcription factor-kappaB through AT1 and AT2 receptors
1
2002
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
A subset of five human mitochondrial formyl peptides mimics bacterial peptides and functionally deactivates human neutrophils
1
2018
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Oxidized mitochondrial DNA activates the NLRP3 inflammasome during apoptosis
1
2012
... In addition, there are many other ways of induction of the main proinflammatory transcription factor NF-kB: overeating, obesity [34], dysbiosis [35], psychological and chronic stress [36], vitamin D deficiency [37], circadian rhythm disturbance [38], aldosterone [39], angiotensin II [40], mitochondrial N-formyl peptides [41], oxidized mitochondrial DNA [42] (Fig. 5). ...
Influence of non-steroidal anti-inflammatory drugs on Drosophila melanogaster longevity
1
2015
... In our experiments on the Drosophila model, we inhibited the different underlying pathways of NF-kB. In most cases, this led to an increase in lifespan [43-45]. ...
Selective anticancer agents suppress aging in Drosophila
0
2013
Pharmacological inhibition of NF-kappaB prolongs lifespan of Drosophila melanogaster
1
2011
... In our experiments on the Drosophila model, we inhibited the different underlying pathways of NF-kB. In most cases, this led to an increase in lifespan [43-45]. ...
Plant polyphenols as inhibitors of NF-kappaB induced cytokine production-a potential anti-inflammatory treatment for Alzheimer's disease?
1
2015
... It is worth noting that many substances of natural origin contained in food can inhibit NF-kB [46]. ...
The Gut Microbiota and Healthy Aging: A Mini-Review
1
2018
... Aging is accompanied by gut microbiota alteration, like decreased overall diversity and an increased abundance of proinflammatory species, that can be a part of systemic inflammation and many aging-related diseases [47, 48]. ...
Gut Microbiota and Extreme Longevity
1
2016
... Aging is accompanied by gut microbiota alteration, like decreased overall diversity and an increased abundance of proinflammatory species, that can be a part of systemic inflammation and many aging-related diseases [47, 48]. ...
Editorial: Immunology of Aging
1
2019
... Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55]. ...
CD28(null) pro-atherogenic CD4 T-cells explain the link between CMV infection and an increased risk of cardiovascular death
1
2018
... Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55]. ...
HPV prevalence and HPV-related dysplasia in elderly women
1
2018
... Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55]. ...
A 3D human brain-like tissue model of herpes-induced Alzheimer's disease
1
2020
... Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55]. ...
Interaction between APOE4 and herpes simplex virus type 1 in Alzheimer's disease
0
2020
Herpes simplex virus type 1 and Alzheimer's disease: link and potential impact on treatment
0
2019
Multiscale Analysis of Independent Alzheimer's Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks by Human Herpesvirus
1
2018
... Internal virome aging-related changes can affect different age-dependent diseases, including immune-senescence [49] and atherosclerosis [50] by cytomegalovirus, cancerogenesis by papillomaviruses [51] and Alzheimer’s by simple herpes [52-55]. ...
Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention
2
2019
... Immune parameters associated with survival may vary in diverse populations of different ages. Therefore, we have to focus on the changes considered to be the hallmarks of immunosenescence, based on the literature data. The hallmarks of immunosenescence include: (i) a reduced ability to respond to new antigens; (ii) the accumulation of memory T cells; (iii) a lingering level of low-grade inflammation termed “inflamm-aging.” Mechanistically, immunosenescence is only partially explained by organismal and cellular senescence. Therefore, these hallmarks of immunosenescence would be markedly affected by the history of individual exposure to pathogens. In fact, several factors, such as genetics, nutrition, exercise, previous exposure to microorganisms, biological and cultural sex, and human cytomegalovirus (HCMV) status can influence immunosenescence [56]. ...
... Changes occurring during aging (reproduced with permission from [56]). ...
Sex, gender and immunosenescence: a key to understand the different lifespan between men and women?
1
2013
... Concerning sex/gender, in Western countries, women live 5-6 years more than men do. Furthermore, 85% of over 100 years old are women. It is debated whether women live longer than men for reasons of gender or sex, e.g., for cultural or biological differences. However, females live longer than males in other animal species. There is sexual dimorphism in the immune response, i.e. females are more resistant to infections, but they have a higher incidence of autoimmune diseases compared to males, yet their relevance for life span is negligible. However, age-related changes in various immunological parameters differ between men and women. Findings indicate that the slower rate of decline in immunological parameters in women than those in men is consistent with the fact that women live longer, than men do [57]. ...
Role of Immunogenetics in the Outcome of HCMV Infection: Implications for Ageing
1
2019
... Concerning HCMV, virus status has a greater impact than age on the immune system because the virus contributes to shaping the immune profile and function during normal human aging. HCMV seropositivity is closely related to the reversal of the CD4/CD 8 T-cell ratio. In fact, persistent HCMV infection leads to chronic stimulation of CD8 T cells, which expand clonally showing an effector memory phenotype characterized by low CD28 expression. The absolute increase in memory T cells, called memory inflation, is observed only in older people infected by HCMV [58]. ...
Thymic fatness and approaches to enhance thymopoietic fitness in aging
1
2010
... One of the pillars of adaptive immunity is the thymus. After an active period of creation and training of new T cells in childhood, at the time of puberty, the thymus undergoes involution, losing the stromal part and filling with fat [59], that can decrease T-cell repertoire to new antigens, including SARS-CoV-2. The involution is continuing during aging, because Wnt4 expression is down-regulated, while their Frizzled receptors and PPARgamma expression increases in the thymus [60]. On the contrary, peripheral T-cell numbers are maintained through the antigen-independent homeostatic proliferation of naive T cells that may lead to the emergence of dysfunctional memory-phenotype CD4+ T cell subpopulation (cell senescence-associated T cells, SA-T cells) [61]. SA-T cells secrete abundant pro-inflammatory factors such as osteopontin and chemokines, playing a direct role in SASP [62]. ...
Transgenic Exosomes for Thymus Regeneration
1
2019
... One of the pillars of adaptive immunity is the thymus. After an active period of creation and training of new T cells in childhood, at the time of puberty, the thymus undergoes involution, losing the stromal part and filling with fat [59], that can decrease T-cell repertoire to new antigens, including SARS-CoV-2. The involution is continuing during aging, because Wnt4 expression is down-regulated, while their Frizzled receptors and PPARgamma expression increases in the thymus [60]. On the contrary, peripheral T-cell numbers are maintained through the antigen-independent homeostatic proliferation of naive T cells that may lead to the emergence of dysfunctional memory-phenotype CD4+ T cell subpopulation (cell senescence-associated T cells, SA-T cells) [61]. SA-T cells secrete abundant pro-inflammatory factors such as osteopontin and chemokines, playing a direct role in SASP [62]. ...
The impact of senescence-associated T cells on immunosenescence and age-related disorders
1
2018
... One of the pillars of adaptive immunity is the thymus. After an active period of creation and training of new T cells in childhood, at the time of puberty, the thymus undergoes involution, losing the stromal part and filling with fat [59], that can decrease T-cell repertoire to new antigens, including SARS-CoV-2. The involution is continuing during aging, because Wnt4 expression is down-regulated, while their Frizzled receptors and PPARgamma expression increases in the thymus [60]. On the contrary, peripheral T-cell numbers are maintained through the antigen-independent homeostatic proliferation of naive T cells that may lead to the emergence of dysfunctional memory-phenotype CD4+ T cell subpopulation (cell senescence-associated T cells, SA-T cells) [61]. SA-T cells secrete abundant pro-inflammatory factors such as osteopontin and chemokines, playing a direct role in SASP [62]. ...
Physiology and pathology of T-cell aging
1
2020
... One of the pillars of adaptive immunity is the thymus. After an active period of creation and training of new T cells in childhood, at the time of puberty, the thymus undergoes involution, losing the stromal part and filling with fat [59], that can decrease T-cell repertoire to new antigens, including SARS-CoV-2. The involution is continuing during aging, because Wnt4 expression is down-regulated, while their Frizzled receptors and PPARgamma expression increases in the thymus [60]. On the contrary, peripheral T-cell numbers are maintained through the antigen-independent homeostatic proliferation of naive T cells that may lead to the emergence of dysfunctional memory-phenotype CD4+ T cell subpopulation (cell senescence-associated T cells, SA-T cells) [61]. SA-T cells secrete abundant pro-inflammatory factors such as osteopontin and chemokines, playing a direct role in SASP [62]. ...
The CD8 T Cell Response to Respiratory Virus Infections
1
2018
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
Natural killer cell responses during viral infections: flexibility and conditioning of innate immunity by experience
1
2011
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
The role of CD8+ T-cell replicative senescence in human aging
1
2005
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
Immune Clearance of Senescent Cells to Combat Ageing and Chronic Diseases
1
2020
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
Senescent cells evade immune clearance via HLA-E-mediated NK and CD8+ T cell inhibition
1
2019
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
Contributions of Age-Related Thymic Involution to Immunosenescence and Inflammaging
1
2020
... CD8+ cytotoxic T lymphocytes [63] and NK cells [64] clear cells infected by viruses (e.g. SARS-CoV-2). However, CD8+ themselves undergo senescence [65]. Immunosenescence could be the reason for the dysfunction of immune clearance of senescent cells [66]. In addition, senescent cells avoid immune clearance through HLA-E-mediated inhibition of NK and CD8 + T cells [67]. Thus, thymus involution is one of the mechanisms of inflamm-aging [68] (Fig. 6). ...
Remodeling of Bone Marrow Hematopoietic Stem Cell Niches Promotes Myeloid Cell Expansion during Premature or Physiological Aging
1
2019
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
Aging of hematopoietic stem cells
1
2018
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
Extracellular Matrix Regulation of Stem Cell Behavior
1
2016
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
Inflammation: a key regulator of hematopoietic stem cell fate in health and disease
1
2017
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
Signaling Pathways Regulating Hematopoietic Stem Cell and Progenitor Aging
1
2018
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
The hematopoietic stem cell niche: from embryo to adult
1
2018
... Senescence of bone marrow hematopoietic stem cells is affected by (HSC) niche [69] and intrinsic factors [70], extracellular matrix stiffness [71], systemic inflammation [72] or other systemic factors [73]. With age, HSCs reduce the homing and regenerative capacity and increase proinflammatory myeloid-biased differentiation [74]. ...
The emerging role of T follicular helper (TFH) cells in aging: Influence on the immune frailty
1
2020
... T follicular helper (TFH) cells are presented in lymphoid organs and in peripheral blood and help B cells for the production of immunoglobulins. Dysfunctional TFH cells with aging play a role in cancer, autoimmune and cardiovascular diseases [75]. ...
Single-cell transcriptomics of the naked mole-rat reveals unexpected features of mammalian immunity
1
2019
... Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80]. ...
Genome analysis reveals insights into physiology and longevity of the Brandt's bat Myotis brandtii
1
2013
... Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80]. ...
... Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80]. ...
Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host
1
2019
... Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80]. ...
Insights into the evolution of longevity from the bowhead whale genome
1
2015
... Recent studies revealed that long-lived mammalian species are characterized by the particularities in their immune system. Cancer and other age-related disease-resistant naked mole-rats lack canonical natural killer cells [76]. Many expanded gene families in the longest-living microbat Myotis brandti are involved in the immune response [77]. Bats showed a unique, age-related pattern of gene expression associated with DNA repair, autophagy, immunity and tumor suppression, which can lead to an increase in their health span [78]. They also express a reduced inflammation response after viral infection [79]. The evaluation of the bowhead whale genome revealed the potentially relevant changes in genes related to the immune response [80]. ...
Immune parameters identify Italian centenarians with a longer five-year survival independent of their health and functional status
1
2014
... Human centenarians are a model for healthy aging. The longest living cohort of Italian centenarians has more favorable values of important immune parameters: naïve, activated/memory and effector/memory T cells [81]. Healthy centenarians presented with a distinct expression of proteins/pathways that reflect a healthy immune function, including less inflamm-aging and autoimmunity and increased B cell-mediated immune response [82]. ...
Successful aging: insights from proteome analyses of healthy centenarians
1
2020
... Human centenarians are a model for healthy aging. The longest living cohort of Italian centenarians has more favorable values of important immune parameters: naïve, activated/memory and effector/memory T cells [81]. Healthy centenarians presented with a distinct expression of proteins/pathways that reflect a healthy immune function, including less inflamm-aging and autoimmunity and increased B cell-mediated immune response [82]. ...
Sicilian centenarian offspring are more resistant to immune ageing
2
2019
... Centenarians have been used as an optimal model for successful aging. However, this model shows several limitations, in particular the selection of appropriate controls. Thus, the interest has been centered on centenarian offspring, since it is well known that they are healthier than the remaining old people are. Accordingly, significant differences between old subjects and centenarian offspring, in most of the studied T and B subsets, show that centenarian offspring subsets present intermediate phenotyping between old and younger people. Therefore, centenarian offspring retain more youthful immunological parameters and the exhaustion of the immune system is less evident than in old people without centenarian parents [83]. Therefore, cell subset changes could represent a hallmark of successful or unsuccessful aging and could be used as a biomarker of human life span, potentially useful for the evaluation of immunosenescence treatment [83, 84]. ...
... ]. Therefore, cell subset changes could represent a hallmark of successful or unsuccessful aging and could be used as a biomarker of human life span, potentially useful for the evaluation of immunosenescence treatment [83, 84]. ...
B cells compartment in centenarian offspring and old people
1
2010
... Centenarians have been used as an optimal model for successful aging. However, this model shows several limitations, in particular the selection of appropriate controls. Thus, the interest has been centered on centenarian offspring, since it is well known that they are healthier than the remaining old people are. Accordingly, significant differences between old subjects and centenarian offspring, in most of the studied T and B subsets, show that centenarian offspring subsets present intermediate phenotyping between old and younger people. Therefore, centenarian offspring retain more youthful immunological parameters and the exhaustion of the immune system is less evident than in old people without centenarian parents [83]. Therefore, cell subset changes could represent a hallmark of successful or unsuccessful aging and could be used as a biomarker of human life span, potentially useful for the evaluation of immunosenescence treatment [83, 84]. ...
Is anti-ageing drug discovery becoming a reality?
2
2020
... Based on the present level of knowledge, the geroprotective therapies targeting the mechanisms of immunosenescence are just emerging [85]. Their studies need to intensify, with a broader identification of potential clinically applicable interventions and biomarkers, and their extensive pre-clinical and clinical testing [85]. ...
... ]. Their studies need to intensify, with a broader identification of potential clinically applicable interventions and biomarkers, and their extensive pre-clinical and clinical testing [85]. ...
Geroprotectors.org: a new, structured and curated database of current therapeutic interventions in aging and age-related disease
1
2015
... There are several hundreds of potential geroprotective interventions, that have been demonstrated on model organisms and collected in online databases Geroprotectors.Org [86] and DrugAge [87]. Not all of them meet the criteria of effective and safe treatment applicable for humans [88, 89]. ...
The DrugAge database of aging-related drugs
1
2017
... There are several hundreds of potential geroprotective interventions, that have been demonstrated on model organisms and collected in online databases Geroprotectors.Org [86] and DrugAge [87]. Not all of them meet the criteria of effective and safe treatment applicable for humans [88, 89]. ...
Developing criteria for evaluation of geroprotectors as a key stage toward translation to the clinic
1
2016
... There are several hundreds of potential geroprotective interventions, that have been demonstrated on model organisms and collected in online databases Geroprotectors.Org [86] and DrugAge [87]. Not all of them meet the criteria of effective and safe treatment applicable for humans [88, 89]. ...
Geroprotectors: A Unified Concept and Screening Approaches
1
2017
... There are several hundreds of potential geroprotective interventions, that have been demonstrated on model organisms and collected in online databases Geroprotectors.Org [86] and DrugAge [87]. Not all of them meet the criteria of effective and safe treatment applicable for humans [88, 89]. ...
The road ahead for health and lifespan interventions
1
2020
... According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94]. ...
Debunking the Myth of Exercise-Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health Across the Lifespan
1
2018
... According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94]. ...
Prolonged fasting reduces IGF-1/PKA to promote hematopoietic-stem-cell-based regeneration and reverse immunosuppression
1
2014
... According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94]. ...
When Fasting Gets Tough, the Tough Immune Cells Get Going-or Die
1
2019
... According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94]. ...
Influence of caloric restriction on aging immune system
1
2004
... According to the clinical studies conducted around the world, exercise, fasting, caloric restriction, resveratrol, metformin and NAD precursors are the interventions with the highest number of clinical trials that target aging [90]. For all of them, a geroprotective effect on immunity is shown. Epidemiological data indicate that regular physical activity reduces the incidence of infectious diseases in the elderly, including viral and bacterial infections, as well as non-infectious diseases associated with the immune system, such as cancer and chronic inflammatory diseases [91]. Cyclic fasting decelerated the immunosuppression caused by chemotherapy and reversed age-dependent myeloid-bias in mice [92]. Cycles of fasting reduce autoimmunity and activate the lymphocyte-dependent killing of cancer cells in humans [93]. The immunological status of rodents under calorie restriction is superior to the immunological status of the non-restricted animals, involving activation of the upstream signaling molecules and cytokine gene expression that are altered with age [94]. ...
Long-Term Effects of Dietary Protein and Branched-Chain Amino Acids on Metabolism and Inflammation in Mice
2
2018
... Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
... ]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
Interleukin-5 and IL-5 receptor in health and diseases
1
2011
... Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
Nutritional disorders in the elderly
1
2006
... Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance
1
2005
... Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
Effect of L-alanine and some other amino acids on thymocyte proliferation in vivo
1
1993
... Arguably, there is no need to limit the entire diet. It is enough just to reduce the intake of certain nutrients to the necessary minimum. Protein restriction increased circulating interleukin-5 concentration in mice [95], that experimental overexpression in vivo significantly increases the number of eosinophils and B cells [96]. However, protein undernutrition is unfavorable for immune function in the elderly [97]. Methionine-deficient diet extends mouse lifespan and slows immune aging [98]. Branched-chain amino acid supplementation induced pro-inflammatory gene expression in visceral adipose tissue in mice [95]. On the contrary, treatment by other animo acids may decrease the aging-related loss of immune system function. Thus, alanine supplementation has stimulated the proliferation of immune cells [99]. ...
The quest to slow ageing through drug discovery
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Lithium's antiviral effects: a potential drug for CoViD-19 disease?
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Review of lithium effects on immune cells
1
2015
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
T cells with dysfunctional mitochondria induce multimorbidity and premature senescence
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Metformin Enhances Autophagy and Normalizes Mitochondrial Function to Alleviate Aging-Associated Inflammation
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Effects of Acarbose on the Gut Microbiota of Prediabetic Patients: A Randomized, Double-blind, Controlled Crossover Trial
1
2017
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Effects of acarbose and metformin on the inflammatory state in newly diagnosed type 2 diabetes patients: a one-year randomized clinical study
1
2019
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Influence of Resveratrol on the Immune Response
1
2019
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Polyamines reverse immune senescence via the translational control of autophagy
1
2020
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Senescent cell clearance by the immune system: Emerging therapeutic opportunities
1
2018
... Brian Kennedy et al. reviewed potential geroprotectors and paid special attention to rapamycin, senolytics, metformin, acarbose, spermidine, NAD+ enhancers and lithium [100]. Lithium presents a clear antiviral activity demonstrated at the preclinical level [101]. Lithium chloride confers protection against viral myocarditis via suppression of coxsackievirus B3 virus replication [102]. Lithium affects many aspects of immunity, including the activity of B- and T-cells, macrophages, interleukin-2 levels [103]. NAD precursors alleviate dysfunctional mitochondria in T cells [104]. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation [105]. Acarbose benefits for immune function may be mediated by selective modulation of the gut microbiota [106]. After one year of treatment with acarbose or metformin, IL-6, TNF-α, IL-1β and ferritin levels of pro-inflammatory factors in type 2 diabetes patients were significantly decreased [107]. In accordance with a review [108], resveratrol can suppress the toll-like receptor and pro-inflammatory genes’ expression, associated with widespread health benefits for different autoimmune and chronic inflammatory diseases. Spermidine induces autophagy and improves the function of both the old mouse and old human B cells [109]. It has been proposed, that senolytics, i.e. drugs that selectively eliminate senescent cells that are the main source of pro-inflammatory cytokines with aging, may prove to alleviate immune dysfunction in older individuals [110]. However, this assumption requires experimental confirmation. ...
Superiority of sirolimus (rapamycin) over cyclosporine in augmenting allograft and xenograft survival in mice treated with antilymphocyte serum and donor-specific bone marrow
1
1997
... Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115]. ...
Effects of molecularly targeted therapies on murine thymus: highly selective mTOR inhibitors induce reversible thymic involution
1
2015
... Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115]. ...
A randomized control trial to establish the feasibility and safety of rapamycin treatment in an older human cohort: Immunological, physical performance, and cognitive effects
1
2018
... Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115]. ...
Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2
1
2020
... Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115]. ...
Melatonin and its influence on immune system
1
2007
... Rapamycin is a well-known potent immune-suppressive agent in xenotransplantation [111]. Rapamycin caused reversible thymus involution in mice [112]. Nonetheless, in a randomized control trial in an older human cohort, rapamycin increased a myeloid cell subset and TREGS [113]. Network-based transcriptomic drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2 revealed rapamycin along with melatonin and mercaptopurine as potential anti-HCoV drugs [114]. A large number of reports have documented a relationship between melatonin and the immune system [115]. ...

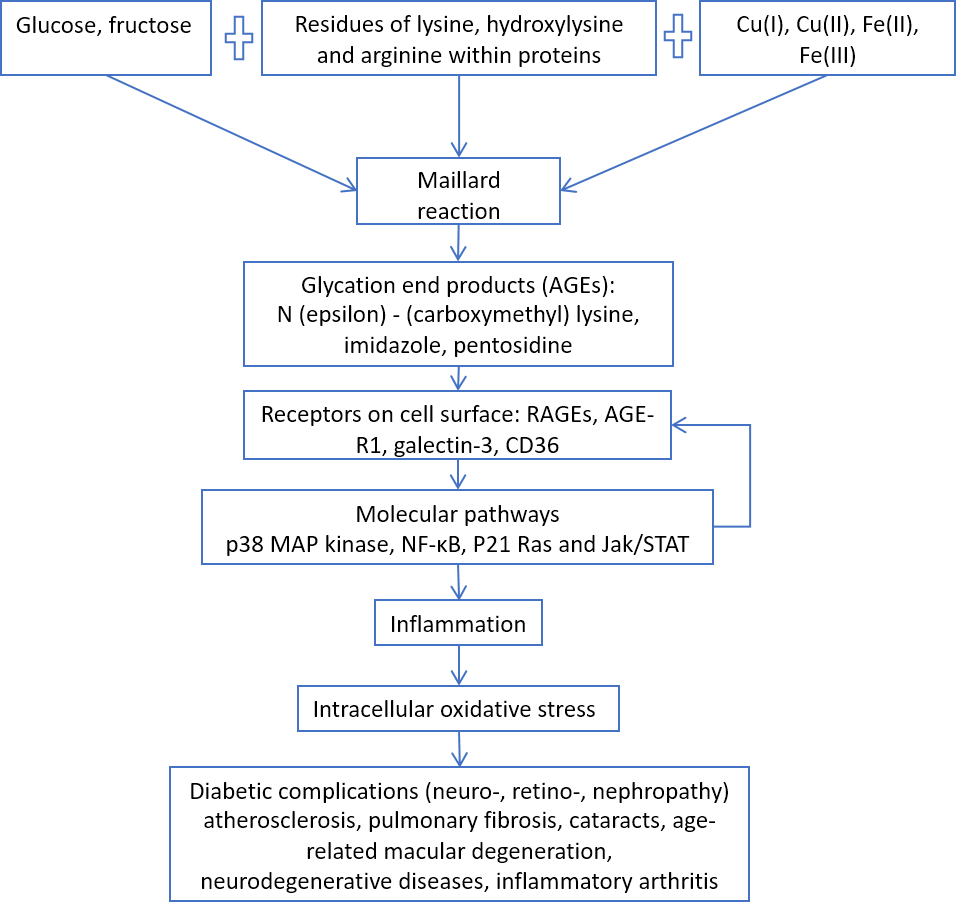
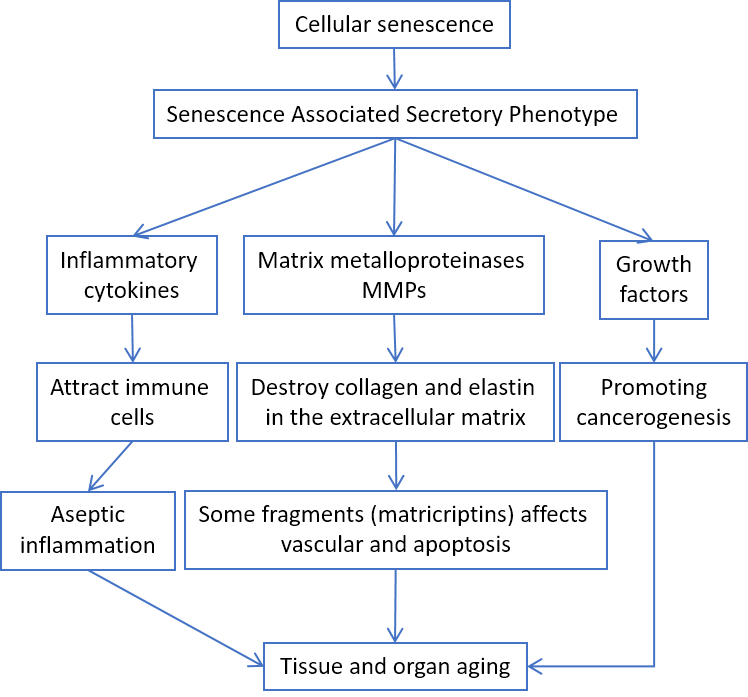
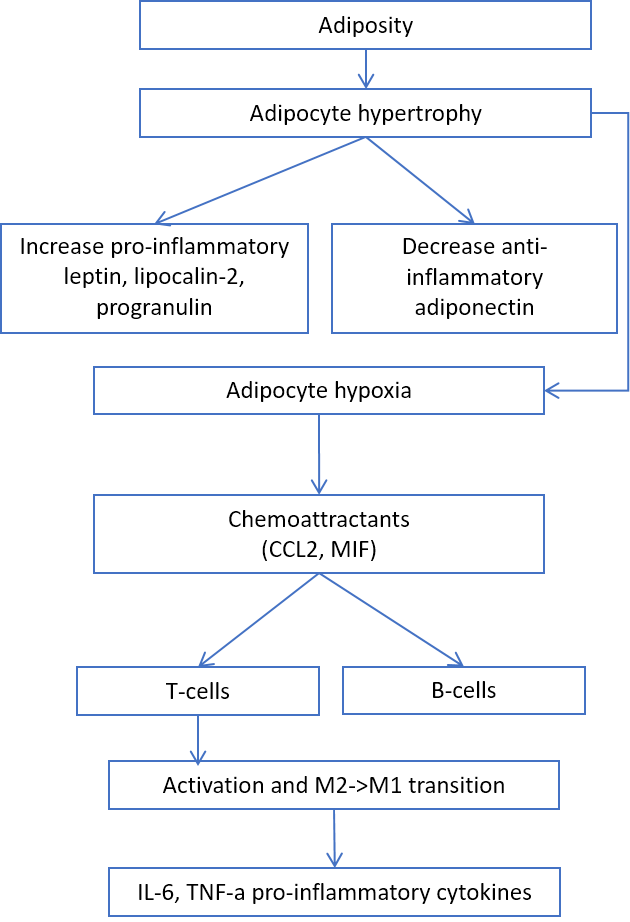
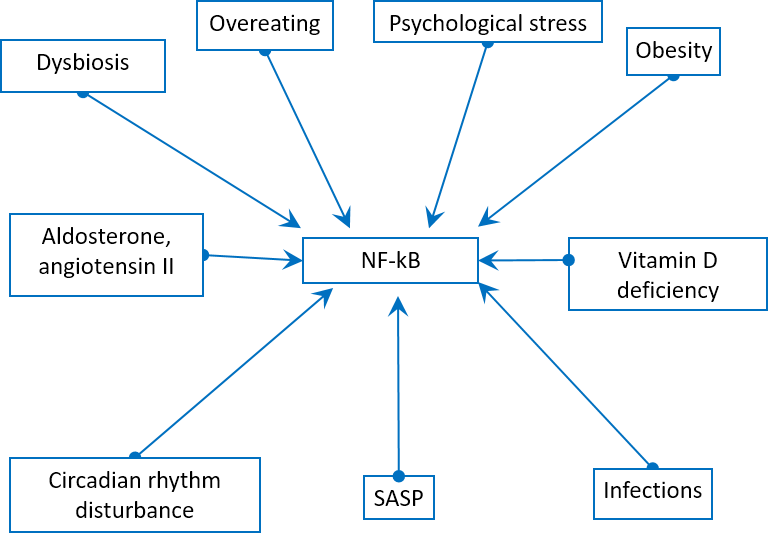
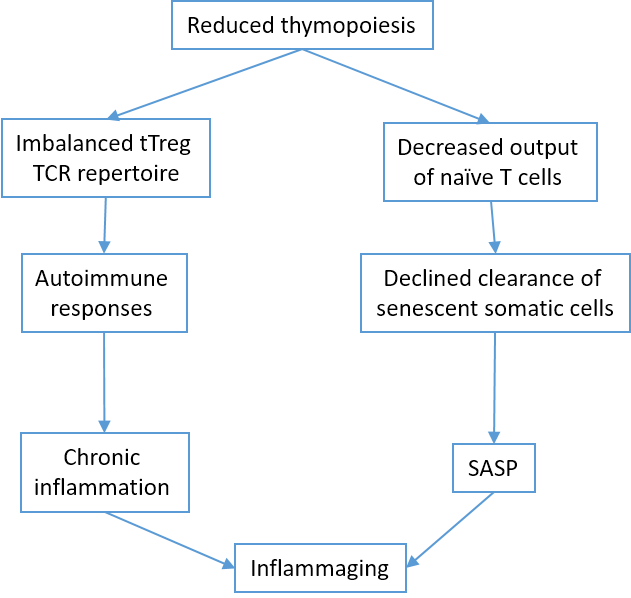
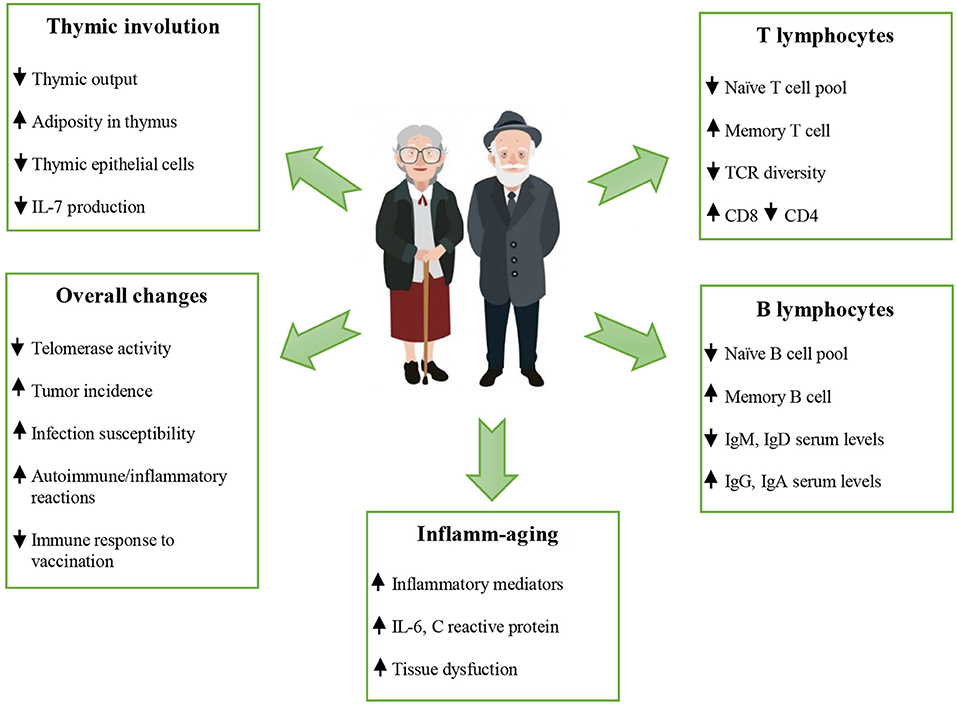
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



