Oxidative Stress at the Crossroads of Aging, Stroke and Depression
Shao Anwen1, *, #, Lin Danfeng2, #, Wang Lingling2, Tu Sheng3, Lenahan Cameron4, 5, Zhang Jianmin1, 6, 7
1Department of Neurosurgery, Second Affiliated Hospital, School of Medicine, Zhejiang University, Zhejiang, China.2Department of Surgical Oncology, Second Affiliated Hospital, School of Medicine, Zhejiang University, Zhejiang, China.3State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital, College of Medicine, Zhejiang University, Zhejiang, China.4Burrell College of Osteopathic Medicine, Las Cruces, USA.5Center for Neuroscience Research, School of Medicine, Loma Linda University, Loma Linda, CA, USA.6Brain Research Institute, Zhejiang University, Zhejiang, China.7Collaborative Innovation Center for Brain Science, Zhejiang University, Zhejiang, China.
通讯作者: Correspondence should be addressed to: Dr. Anwen Shao, Second Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou, Zhejiang, China. Email: 21118116@zju.edu.cn; anwenshao@sina.com;Correspondence should be addressed to: Dr. Anwen Shao, Second Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou, Zhejiang, China. Email: 21118116@zju.edu.cn; anwenshao@sina.com;Correspondence should be addressed to: Dr. Anwen Shao, Second Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou, Zhejiang, China. Email: 21118116@zju.edu.cn; anwenshao@sina.com;
收稿日期:2020-01-28
修回日期: 2020-02-18
接受日期: 2020-02-25
网络出版日期: 2020-12-01
版权声明:
2020 this is an open access article distributed under the terms of the creative commons attribution license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
作者简介:
These authors contributed equally to this work.
展开
Abstract
Epidemiologic studies have shown that in the aging society, a person dies from stroke every 3 minutes and 42 seconds, and vast numbers of people experience depression around the globe. The high prevalence and disability rates of stroke and depression introduce enormous challenges to public health. Accumulating evidence reveals that stroke is tightly associated with depression, and both diseases are linked to oxidative stress (OS). This review summarizes the mechanisms of OS and OS-mediated pathological processes, such as inflammation, apoptosis, and the microbial-gut-brain axis in stroke and depression. Pathological changes can lead to neuronal cell death, neurological deficits, and brain injury through DNA damage and the oxidation of lipids and proteins, which exacerbate the development of these two disorders. Additionally, aging accelerates the progression of stroke and depression by overactive OS and reduced antioxidant defenses. This review also discusses the efficacy and safety of several antioxidants and antidepressants in stroke and depression. Herein, we propose a crosstalk between OS, aging, stroke, and depression, and provide potential therapeutic strategies for the treatment of stroke and depression.
ShaoAnwen, LinDanfeng, WangLingling, TuSheng, LenahanCameron, ZhangJianmin. Oxidative Stress at the Crossroads of Aging, Stroke and Depression[J]. Aging and Disease, 2020, 11(6): 1537-1566 https://doi.org/10.14336/AD.2020.0225
1. Introduction
Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11].
Over the past two decades, studies have identified the role of OS in these two diseases. Recently, preclinical experiments and clinical trials have focused on studying the efficacy of antioxidants and combined therapy with antidepressants in stroke or depressed patients. We review the results of experiments within the last 5 years, providing a comprehensive and novel overview of this interesting field.
Figure 1. Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303].
2. Overview of oxidative stress and the antioxidant system
OS describes a state in which the body produces excessive ROS and RNS in response to deleterious substances. Under normal conditions, OS and the anti-OS system are beneficial to physiological functions, such as respiration, circulation, etc. Mitochondrial dysfunction and internal environment disturbance can arise from multiple cellular molecules and signaling pathways as a result of imbalanced redox reactions, characterized by increased production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Fig. 1), as well as decreased antioxidant defenses. The most important ROS are mitochondrial ROS (mitoROS) produced from enzymes of the mitochondrial respiratory chain. Overproduced mitoROS can affect metabolic pathways, leading to a compromised function of organelles [12]. Another significant source of ROS is NADPH oxidase (NOX). When the complex of NOX is assembled, it transports an electron from the cytosolic NADPH to oxygen to then form superoxide on the extracellular side [13]. The antioxidant system consists of enzymes and nonenzymatic substances, among which nuclear factor erythroid 2-related factor (Nrf2) is the most important factor. Nrf2 is a transcription factor and can recognize the antioxidant response element (ARE) to regulate several genes, such as heme oxygenase 1 (HO-1). Activation of the Nrf2-ARE pathway increases antioxidants and protects the cells from free radical (FR) accumulation [14].
Under physiological conditions, moderate OS activity is necessary for body health. Toxic effects derived from ROS and RNS can be ameliorated or neutralized by FR scavengers and the antioxidant system. However, when a large number of ROS and RNS are generated, excessive FR then induce molecular oxidation, cell membrane modification, and enzyme inactivation, resulting in cellular damage and functional decline [15]. Overactive OS with an imbalanced redox state can induce many injuries, especially to the brain, partly expounding the role of OS in both stroke and depression.
3. Overview of stroke and depression
Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20].
In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS.
As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2).
4. Roles of oxidative stress in stroke and depression
Following ischemic stroke, an excess of ROS and RNS causes an ascending concentration of H+ and H2O2, leading to DNA damage, endothelial impairment, and mitochondrial dysfunction (see below for details). Furthermore, they exacerbate the damage through OS-mediated inflammation, apoptosis, autophagy, and the microbial-gut-brain axis. Apart from the mechanisms shown in ischemic stroke, the ROS and RNS, derived from the hemoglobin-heme-iron axis and activation of NOX, increase glutamate and inflammatory activities in ICH, and they disrupt BBB integrity to influence EBI and SBI in SAH. It is unsurprising to find semblable outcomes and pathological processes of OS in depression.
Figure 2. Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37].
4.1 Oxidative stress in stroke and depression
Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45].
Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53].
Figure 3. Mechanism of oxidative stress in ischemic stroke.
In SAH, EBI is identified as the immediate injury accompanied by subsequent events (brain edema, inflammation, apoptosis, etc.) in the first 72 hours, and EBI plays a vital role in the pathological processes (Fig. 4). After the occurrence of SAH, the intracranial pressure will rise to the pressure of the diastolic artery, which reduces cerebrospinal fluid pressure and disturbs cerebral autoregulation before BBB breakdown and occurrence of cerebral edema [54]. Due to erythrocytosis, the central nervous system (CNS) is exposed to high levels of Hb and Hb-degradation products in the subarachnoid space. This pathological process produces excess ROS and RNS, and promotes cerebral vasospasm, cerebrovascular stenosis, and delayed cerebral ischemia [55]. Furthermore, several harmful events occur in SAH survivors, including altered ionic homeostasis, excitotoxicity, destruction of vascular integrity, OS, inflammation, apoptosis, autophagy, and activation of the NOS pathway [56-62].
The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75].
4.2 Common oxidative stress-mediated process between stroke and depression
In previous sections, it is mentioned that OS-mediated pathological mechanisms (e.g., inflammation and apoptosis) act as a bridge and exert functions in stroke and depression (Fig. 5). Details about common OS-induced mechanisms are shown as follows.
4.2.1 Oxidative stress-mediated inflammation
Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56].
Figure 4. Schematic model of oxidative mechanisms in ICH and SAH, especially associated with hemoglobin (Hb). During the hemoglobin-heme-iron axis, Hb is released into the extracellular space and is accompanied by an abundance of superoxide generated from the non-enzymatic oxidation of Hb [46]. This oxidation of Hb produces methemoglobin, which releases heme to stimulate lipid peroxidation and other oxidative actions around the hematoma in brain tissue. Meanwhile, iron released from Hb degradation is used in the Fenton reaction to transform H2O2 into the hydroxyl radical, leading to increased oxidative damage [47].
In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90].
Figure 5. Mechanisms of oxidative stress and OS-mediated cell death pathway in depression.
4.2.2 Oxidative stress-mediated apoptosis
Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60].
Parallel to OS-mediated apoptosis in stroke, intrinsic apoptosis associated with mitochondria occupies an important position in the development of depression. When exposed to chronic stress, overactive apoptosis stimulated by OS in neurons promotes the neurodegeneration in depressed animals [97]. Mechanistically, Bcl-2-mediated damage and excess levels of Ca2+ trigger various kinases, leading to a loss of the mitochondrial membrane potential, and an increased release of intermembrane proteins into the cytoplasm. In the presence of dATP, the released Cytc binds to the cytosolic Apaf 1 to activate caspase-9 and promote apoptosome formation. Meanwhile, enhanced permeability of the outer membrane of the mitochondria allows apoptosis-inducing factor (AIF) and endonuclease G to move to the nucleus and regulate chromatinolysis [97]. In addition, chronic mild stress suppresses the expression of Bcl-2-associated athanogene-1, a gene involved in anti-apoptosis, resulting in the activation of caspases, BAX, and Bcl-2 antagonists in the mitochondria. Consequently, there is an excess of neuronal death [98]. Moreover, it is highlighted that polymorphisms of apoptotic protein genes may be associated with MDD [99].
Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed.
In depression, the changes of microbiota, intestinal composition, and the resultant bacterial translocation expand the content of two-way interaction of the gut-brain axis. Zhang P et al. recognized the gut microbiome as an environmental factor that can shape the brain through this axis. Data from clinical sampling showed a significant difference of gut microbiota compositions between MDD patients and healthy controls, demonstrating that gut dysbiosis may mediate the host's metabolism to promote depressive-like behaviors [109]. In addition, gut dysbiosis could have an impact on the biology of MDD through several pathways involving immune activation, ROS and RNS, and neuroplasticity cascades [110]. Therefore, the microbiota-gut-brain axis may play a key role in the pathophysiological process of MDD. This axis opens a new preventative and therapeutic perspective for depression. However, due to methodological inconsistencies and limitations, more fundamental and clinical research should be undertaken.
5. Roles of aging in stroke and depression via oxidatvie stress
The preceding content shows us a close connection between stroke and depression via the OS system and OS-mediated biological processes. Moreover, numerous studies have proved that aging or senescence is a risk factor that aggravates stroke and depression. For example, the population-based multiethnic cohort reported that the risk effect of physical inactivity is modified by age, and there is a conspicuous risk found only in stroke patients >80 years of age [111, 112]. Population-based surveillance studies predicted that the number of strokes will double by 2050, with incidence occurring primarily in the elderly (>age 75) groups [113]. Although depression prevalence in the elderly varies across studies, Sjoberg L et al. indicated that older adults with poor physical function have a higher prevalence of depression [114]. Herein, there is an “aging-stroke-depression” network and OS seems to be the center of this network. So, what relationship does aging have with oxidative stress, stroke, and depression?
5.1 Aging and oxidative stress
There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121].
5.2 Aging and stroke
Normally, physiological events of aging involve a decline of innate functions, including shortened telomeres, dysregulated hormones, and dampened immune responses. Under pathological conditions, aging stimulates and exacerbates cellular injuries, such as DNA damage and FR accumulation, to increase the vulnerability of the brain [122]. Mattson MP et al. depicted ten hallmarks of brain aging (e.g., OS and impaired DNA repair), among which, dysregulated energy metabolism is the core factor [122]. When exposed to ischemia, aging impairs the integrity of the neurovascular unit and damages brain tissues [123]. Aging can also destroy collateral circulation and revascularization of the brain through increased FR and inflammatory responses to aggravate stroke [124]. Age-related cognitive deteriorations also exist in the ICH mice model. Researchers showed a delayed neurological improvement and decreased levels of antioxidants in senescence-accelerated mice than senescence-accelerated resistant mice, though the time period in which the neurological deficits occurred and increased remains the same. To conclude, these results implied a key role of OS in senescence and stroke.
Apart from OS, several other mechanisms are involved in OS and stroke. The ubiquitin proteasome pathway protects neurons by removing abnormal or toxic proteins located in the axons and dendrites, and ischemia may exacerbate injury of an aged brain through impairing ubiquitin proteasome function [125]. Furthermore, aging is detrimental to ischemic stroke recovery due to its role in altering astrocytic proliferation, inhibiting vascular endothelial growth factor (VEGF) production, and upregulating the release of inflammatory cytokines [126, 127]. What about aging in hemorrhagic stroke? Apart from the direct effects of aging in ICH, senescence may contribute to ICH progression by changing the body physiology and increasing the risk of multiple chronic health conditions and comorbidities, such as hypertension and diabetes [128]. However, the exact mechanism remains unclear. To our knowledge, there is no clear evidence explaining a relation between SAH and aging.
5.3 Aging and depression
In the process of aging, biological changes in depression include mitochondrial dysfunction, dopamine dysfunction, and increased proinflammatory cytokines involved in the cellular senescence cycle. In general, mitochondrial abnormalities can be measured in peripheral blood mononuclear cells [129]. For instance, the clinical study by Karabatsiakis et al. depicted higher levels of mitochondrial impairments in older folks with depression when compared with healthy controls [130]. A systematic review and meta-analysis also supported this connection, indicating that late-life depression may be associated with a decreased hippocampal volume, though the relationship was not straightforward [131].
Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link.
5.4 Aging and oxidative stress in stroke or depression
There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders?
Figure 6. Proposed crosslink and interplay among aging, oxidative stress, stroke and depression.
6. Antioxidants and antidepressants in stroke/depression
In the last 5 years, mountains of preclinical and clinical studies have found that the application of antioxidants inhibits stroke and depression in patients with either, or both diseases, although the therapeutic effects and safety are under consideration. What’s more, the main pharmacotherapy for depression patients focuses on antidepressants [143], and the selective serotonin reuptake inhibitors (SSRIs) have been used as first-line antidepressants in recent decades. However, SSRIs have shown moderate efficacy and significant side-effects [10]. Variability in efficacy and acceptability are large in head-to-head trials [144]. Therefore, development of an accurate and effective antidepressant regimen requires our constant efforts. Herein, we list most of the relative studies about antioxidants and antidepressants used in treating stroke/depression, in order to find potential therapies.
6.1 Antioxidants in stroke
Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162].
Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling.
Table 1
Table 1 Antioxidants in Stroke.
Stroke
Antioxidants
Anti-OS activity
Mechanism of anti-OS activity and others
Ischemic stroke
fucoxanthin
anti-OS
inhibit OS via Nrf2/HO-1 signaling pathway
Sirtuin 6
protocatechualdehyde
mangiferin
Korean Red Ginseng
11-Keto-β-boswellic acid
metformin
S-allyl cysteine
monomethyl fumarate
dieckol
fumarate
ursolic acid
upregulate Nrf2 pathway and expression levels of BDNF
HP-1c
AMPK-Nrf2 pathway activation, without any toxicity after penetrating the brain
andrographolide
up-regulate Nrf2/HO-1 expression via regulation of p38 MAPK
Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties.
Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats.
Table 2
Table 2 Antioxidants in Depression.
Antioxidants
Anti-OS activity
Mechanism of anti-OS activity and others
Depression
bay 60-7550
anti-OS
downregulate gp91phox; activate the cAMP/cGMP-pVASP-CREB-BDNF signaling pathway
Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243].
Table 3
Table 3 Co-antioxidants in stroke and depression from experiments.
Clinical trials and outcomes in stroke
Clinical trials and outcomes in depression
Types
Outcomes
Types
Outcomes
flavonoid
meta-analysis
high flavonoid reduces risk of stroke
RCT
higher flavonoid links to lower depression risk especially among women
UA
RCT and URICOICTUS
UA is safe; UA enhances outcomes of stroke
cohort studies and meta-analysis
UA are associated with low risk of depression hospitalization and lower MDA levels
melatonin
RCT
early melatonin usage ameliorates the brain injury of asphyxial newborns
RCT
buspirone-melatonin therapy benefits cognitive function
Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271].
6.3 Co-antioxidants and promising drugs in stroke and depression
Thus far, the use of antioxidants in animals has been successful in treating stroke or depression. According to the previous summarization, we have found some co-antioxidants (Table 3). Additionally, several clinical trials show that co-antioxidants exert protections in stroke and depression, including flavonoid and melatonin.
6.3.1 Flavonoids
In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276].
6.3.2 Uric Acid
UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281].
6.3.3 Melatonin
To our knowledge, few clinical trials have studied the neuroprotective effects of melatonin on cerebral ischemia prevention. A prospective trial involving 45 neonates indicates that it is practicable to provide early administration of melatonin to ameliorate brain injury in a choking newborn infant [282]. In regards to treatment of MDD, preliminary findings of a study reveal that a combination of buspirone with melatonin can partially benefit cognitive function [283].
Table 4
Table 4 Antidepressants in PSD treatment.
Antioxidants in PSD
Clinical trials
Outcomes
fluoxetine
FOCUS
not support routine use of fluoxetine in preventing PSD or promoting function recovery
fluoxetine/paroxetine
meta-analysis of 12 trials
fluoxetine is the worst choice for PSD treatment; paroxetine is the best drug in terms of efficacy and acceptability
meta-analysis of 20 RCTs
citalopram has similar efficacy and safety as other SSRIs but acts faster than them
fluoxetine
FLAME
exhibit a positive connection between motor recovery
escitalopram
Cochrane review
escitalopram is the best tolerated SSRI, followed by sertraline and paroxetine for PSD
escitalopram
RCT
not take effects on depressive symptoms; diarrhea is more likely to occur
escitalopram
RCT
effective at decreasing the incidence of depression in nondepressed patients
Citalopram
RCT
safe for patients with acute ischemic stroke
Citalopram
RCT
different effects in different stages of PSD
citalopram
RCT
SSRI treatment is well tolerated and beneficial in PSD
SSRI
registry-based score-matched follow-up study
pre-stroke SSRI use increases risk of the hemorrhagic stroke; no increased stroke severity and mortality ischemic stroke
milnacipran
RCT
milnacipran prevents post-stroke depression; safe to use without serious adverse events
Although many co-antioxidants are found in animal models of both diseases, only a few clinical trials have confirmed the efficacy of antioxidants in stroke and depression. The difficulty of antioxidants acting through the BBB in patients may be the reason for this result. Nevertheless, OS and OS-mediated pathways are co-mechanisms of stroke and depression. So, what is the role of antidepressants in stroke survivors?
6.3 Antidepressants in post-stroke
At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted.
DISCUSSION
This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases.
Acknowledgments
This work was funded by the China Postdoctoral Science Foundation (2017M612010), National Natural Science Foundation of China (81701144, 81371433, and 8187 0916), and Major Science and Technology project in medical and health of Zhejiang Province (WKJ-ZJ-1615:2016149634). Wish the global epidemic of 2019 novel coronavirus pneumonia ends soon.
Conflict of interest
The authors state that there was no conflict of interest in the preparation of this review.
Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2017: a systematic analysis for the Global Burden of Disease Study 2017
Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980-2017: a systematic analysis for the Global Burden of Disease Study 2017
Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016
ZuoG, ZhangT, HuangL, AraujoC, PengJ, TravisZ, et al. (2019).
Activation of TGR5 with INT-777 attenuates oxidative stress and neuronal apoptosis via cAMP/PKCepsilon/ALDH2 pathway after subarachnoid hemorrhage in rats
SchmaalL, HibarDP, SamannPG, HallGB, BauneBT, JahanshadN, et al. (2017).
Cortical abnormalities in adults and adolescents with major depression based on brain scans from 20 cohorts worldwide in the ENIGMA Major Depressive Disorder Working Group
TsuboiH, SakakibaraH, TatsumiA, Yamakawa-KobayashiK, MatsunagaM, KanekoH, et al. (2019).
Serum IL-6 levels and oxidation rate of LDL cholesterol were related to depressive symptoms independent of omega-3 fatty acids among female hospital and nursing home workers in Japan
WangZ, ZhouF, DouY, TianX, LiuC, LiH, et al. (2018).
Melatonin Alleviates Intracerebral Hemorrhage-Induced Secondary Brain Injury in Rats via Suppressing Apoptosis, Inflammation, Oxidative Stress, DNA Damage, and Mitochondria Injury
VergouwenMD, VermeulenM, van GijnJ, RinkelGJ, WijdicksEF, MuizelaarJP, et al. (2010).
Definition of delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage as an outcome event in clinical trials and observational studies: proposal of a multidisciplinary research group
ZhongJT, YuJ, WangHJ, ShiY, ZhaoTS, HeBX, et al. (2017).
Effects of endoplasmic reticulum stress on the autophagy, apoptosis, and chemotherapy resistance of human breast cancer cells by regulating the PI3K/AKT/mTOR signaling pathway
. Tumour Biol, 39:1010428317697562.
[58]
ShiX, FuY, ZhangS, DingH, ChenJ (2017).
Baicalin Attenuates Subarachnoid Hemorrhagic Brain Injury by Modulating Blood-Brain Barrier Disruption, Inflammation, and Oxidative Damage in Mice
. Oxid Med Cell Longev, 2017:1401790.
[59]
FanLF, HePY, PengYC, DuQH, MaYJ, JinJX, et al. (2017).
Mdivi-1 ameliorates early brain injury after subarachnoid hemorrhage via the suppression of inflammation-related blood-brain barrier disruption and endoplasmic reticulum stress-based apoptosis
DouY, ShenH, FengD, LiH, TianX, ZhangJ, et al. (2017).
Tumor necrosis factor receptor-associated factor 6 participates in early brain injury after subarachnoid hemorrhage in rats through inhibiting autophagy and promoting oxidative stress
. J Neurochem, 142:478-492.
[62]
WuH, NiuH, WuC, LiY, WangK, ZhangJ, et al. (2016).
The autophagy-lysosomal system in subarachnoid haemorrhage
KooYS, KimH, ParkJH, KimMJ, ShinYI, ChoiBT, et al. (2018).
Indoleamine 2,3-Dioxygenase-Dependent Neurotoxic Kynurenine Metabolism Contributes to Poststroke Depression Induced in Mice by Ischemic Stroke along with Spatial Restraint Stress
. Oxid Med Cell Longev, 2018:2413841.
[68]
KudlowP, ChaDS, CarvalhoAF, McIntyreRS (2016).
Nitric Oxide and Major Depressive Disorder: Pathophysiology and Treatment Implications
. Curr Mol Med, 16:206-215.
[69]
RezinGT, CardosoMR, GoncalvesCL, ScainiG, FragaDB, RiegelRE, et al. (2008).
Inhibition of mitochondrial respiratory chain in brain of rats subjected to an experimental model of depression
Oxidative stress and antioxidant parameters in patients with major depressive disorder compared to healthy controls before and after antidepressant treatment: results from a meta-analysis
. J Clin Psychiatry, 76:1658-1667.
[74]
Moreno-FernandezAM, CorderoMD, Garrido-MaraverJ, Alcocer-GomezE, Casas-BarqueroN, Carmona-LopezMI, et al. (2012).
Oral treatment with amitriptyline induces coenzyme Q deficiency and oxidative stress in psychiatric patients
Oxidative Stress and Antioxidant Parameters in Patients With Major Depressive Disorder Compared to Healthy Controls Before and After Antidepressant Treatment: Results From a Meta-Analysis
GotschelF, KernC, LangS, SparnaT, MarkmannC, SchwagerJ, et al. (2008).
Inhibition of GSK3 differentially modulates NF-kappaB, CREB, AP-1 and beta-catenin signaling in hepatocytes, but fails to promote TNF-alpha-induced apoptosis
Targeting glycogen synthase kinase-3 for oxidative stress and neuroinflammation: Opportunities, challenges and future directions for cerebral stroke management
Linking the biological underpinnings of depression: Role of mitochondria interactions with melatonin, inflammation, sirtuins, tryptophan catabolites, DNA repair and oxidative and nitrosative stress, with consequences for classification and cognition
CulmseeC, ZhuC, LandshamerS, BecattiniB, WagnerE, PellecchiaM, et al. (2005).
Apoptosis-inducing factor triggered by poly(ADP-ribose) polymerase and Bid mediates neuronal cell death after oxygen-glucose deprivation and focal cerebral ischemia
SlyepchenkoA, MaesM, JackaFN, KohlerCA, BarichelloT, McIntyreRS, et al. (2017).
Gut Microbiota, Bacterial Translocation, and Interactions with Diet: Pathophysiological Links between Major Depressive Disorder and Non-Communicable Medical Comorbidities
Potentiation of glutathione loss and nerve cell death by the transition metals iron and copper: Implications for age-related neurodegenerative diseases
CiprianiA, FurukawaTA, SalantiG, ChaimaniA, AtkinsonLZ, OgawaY, et al. (2018).
Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: a systematic review and network meta-analysis
Pretreatment with Korean red ginseng or dimethyl fumarate attenuates reactive gliosis and confers sustained neuroprotection against cerebral hypoxic-ischemic damage by an Nrf2-dependent mechanism
Pre-treatment with metformin activates Nrf2 antioxidant pathways and inhibits inflammatory responses through induction of AMPK after transient global cerebral ischemia
CuiYJ, AmarsanaaK, LeeJH, RhimJK, KwonJM, KimSH, et al. (2019).
Neuroprotective mechanisms of dieckol against glutamate toxicity through reactive oxygen species scavenging and nuclear factor-like 2/heme oxygenase-1 pathway
. Korean Journal of Physiology & Pharmacology, 23:121-130.
Nitroxide antioxidant as a potential strategy to attenuate the oxidative/nitrosative stress induced by hydrogen peroxide plus nitric oxide in cultured neurons. Nitric Oxide-Biology and
ChenN, ZhouZ, LiJ, LiB, FengJ, HeD, et al. (2018).
3-n-butylphthalide exerts neuroprotective effects by enhancing anti-oxidation and attenuating mitochondrial dysfunction in an in vitro model of ischemic stroke
Post-ischemic administration of 5-methoxyindole-2-carboxylic acid at the onset of reperfusion affords neuroprotection against stroke injury by preserving mitochondrial function and attenuating oxidative stress
. Biochemical and Biophysical Research Communications, 497:444-450.
ChenZ, MaoX, LiuA, GaoX, ChenX, YeM, et al. (2015).
Osthole, a natural coumarin improves cognitive impairments and BBB dysfunction after transient global brain ischemia in C57 BL/6J mice: involvement of Nrf2 pathway
LiL, YangN, NinL, ZhaoZ, ChenL, YuJ, et al. (2015).
Chinese herbal medicine formula tao hong si wu decoction protects against cerebral ischemia-reperfusion injury via PI3K/Akt and the Nrf2 signaling pathway
ZhouF, WangM, JuJ, WangY, LiuZ, ZhaoX, et al. (2019).
Schizandrin A protects against cerebral ischemia-reperfusion injury by suppressing inflammation and oxidative stress and regulating the AMPK/Nrf2 pathway regulation
Quercetin inhibits LPS-induced adhesion molecule expression and oxidant production in human aortic endothelial cells by p38-mediated Nrf2 activation and antioxidant enzyme induction
WangWM, LiuZ, LiuAJ, WangYX, WangHG, AnD, et al. (2015).
The Zinc Ion Chelating Agent TPEN Attenuates Neuronal Death/apoptosis Caused by Hypoxia/ischemia Via Mediating the Pathophysiological Cascade Including Excitotoxicity, Oxidative Stress, and Inflammation
Inhibition of Myeloperoxidase by N-Acetyl Lysyltyrosylcysteine Amide Reduces Oxidative Stress-Mediated Inflammation, Neuronal Damage, and Neural Stem Cell Injury in a Murine Model of Stroke
YuenCM, YehKH, WallaceCG, ChenKH, LinHS, SungPH, et al. (2017).
EPO-cyclosporine combination therapy reduced brain infarct area in rat after acute ischemic stroke: role of innate immune-inflammatory response, micro-RNAs and MAPK family signaling pathway
TsaiYR, ChangCF, LaiJH, WuJC, ChenYH, KangSJ, et al. (2018).
Pomalidomide Ameliorates H(2)O(2)-Induced Oxidative Stress Injury and Cell Death in Rat Primary Cortical Neuronal Cultures by Inducing Anti-Oxidative and Anti-Apoptosis Effects
WangBD, GuoH, LiX, YueL, LiuHX, ZhaoL, et al. (2018).
Adiponectin Attenuates Oxygen-Glucose Deprivation-Induced Mitochondrial Oxidative Injury and Apoptosis in Hippocampal HT22 Cells via the JAK2/STAT3 Pathway
ShenK, WangY, ZhangY, ZhouH, SongY, CaoZ, et al. (2015).
Cocktail of Four Active Components Derived from Sheng Mai San Inhibits Hydrogen Peroxide-Induced PC12 Cell Apoptosis Linked with the Caspase-3/ROCK1/MLC Pathway
XuXH, LiGL, WangBA, QinY, BaiSR, RongJ, et al. (2015).
Diallyl trisufide protects against oxygen glucose deprivation -induced apoptosis by scavenging free radicals via the PI3K/Akt -mediated Nrf2/HO-1 signaling pathway in B35 neural cells
ZhangK, TuM, GaoW, CaiX, SongF, ChenZ, et al. (2019).
Hollow Prussian Blue Nanozymes Drive Neuroprotection against Ischemic Stroke via Attenuating Oxidative Stress, Counteracting Inflammation, and Suppressing Cell Apoptosis
LiC, WangX, ChengF, DuX, YanJ, ZhaiC, et al. (2019).
Geniposide protects against hypoxia/reperfusion-induced blood-brain barrier impairment by increasing tight junction protein expression and decreasing inflammation, oxidative stress, and apoptosis in an in vitro system
WichaP, TocharusJ, JanyouA, JittiwatJ, ChangtamC, SuksamrarnA, et al. (2017).
Hexahydrocurcumin protects against cerebral ischemia/reperfusion injury, attenuates inflammation, and improves antioxidant defenses in a rat stroke model
Isoquercetin attenuates oxidative stress and neuronal apoptosis after ischemia/reperfusion injury via Nrf2-mediated inhibition of the NOX4/ROS/NF-kappaB pathway
WangCP, ShiYW, TangM, ZhangXC, GuY, LiangXM, et al. (2017).
Isoquercetin Ameliorates Cerebral Impairment in Focal Ischemia Through Anti-Oxidative, Anti-Inflammatory, and Anti-Apoptotic Effects in Primary Culture of Rat Hippocampal Neurons and Hippocampal CA1 Region of Rats
AltermannCDC, SouzaMA, SchimidtHL, IzaguirryAP, MartinsA, GarciaA, et al. (2017).
Short-term green tea supplementation prevents recognition memory deficits and ameliorates hippocampal oxidative stress induced by different stroke models in rats
Green Tea and Red Tea from Camellia sinensis Partially Prevented the Motor Deficits and Striatal Oxidative Damage Induced by Hemorrhagic Stroke in Rats
Dexmedetomidine Protects Against Neurological Dysfunction in a Mouse Intracerebral Hemorrhage Model by Inhibiting Mitochondrial Dysfunction-Derived Oxidative Stress
XuW, LiT, GaoL, LenahanC, ZhengJ, YanJ, et al. (2019).
Sodium Benzoate Attenuates Secondary Brain Injury by Inhibiting Neuronal Apoptosis and Reducing Mitochondria-Mediated Oxidative Stress in a Rat Model of Intracerebral Hemorrhage: Possible Involvement of DJ-1/Akt/IKK/NFkappaB Pathway
XiZ, HuX, ChenX, YangY, RenJ, WangB, et al. (2019).
Protocatechuic acid exerts protective effects via suppression of the P38/JNK- NF-kappaB signalling pathway in an experimental mouse model of intracerebral haemorrhage
KaruppagounderSS, AlimI, KhimSJ, BourassaMW, SleimanSF, JohnR, et al. (2016).
Therapeutic targeting of oxygen-sensing prolyl hydroxylases abrogates ATF4-dependent neuronal death and improves outcomes after brain hemorrhage in several rodent models
Dimethylfumarate alleviates early brain injury and secondary cognitive deficits after experimental subarachnoid hemorrhage via activation of Keap1-Nrf2-ARE system
Nebivolol attenuates cerebral vasospasm both by increasing endothelial nitric oxide and by decreasing oxidative stress in an experimental subarachnoid haemorrhage
LiuH, ZhaoL, YueL, WangB, LiX, GuoH, et al. (2017).
Pterostilbene Attenuates Early Brain Injury Following Subarachnoid Hemorrhage via Inhibition of the NLRP3 Inflammasome and Nox2-Related Oxidative Stress
ZhangT, WuP, ZhangJH, LiY, XuS, WangC, et al. (2018).
Docosahexaenoic Acid Alleviates Oxidative Stress-Based Apoptosis Via Improving Mitochondrial Dynamics in Early Brain Injury After Subarachnoid Hemorrhage
Homocysteine aggravates ROS-induced depression of transmitter release from motor nerve terminals: potential mechanism of peripheral impairment in motor neuron diseases associated with hyperhomocysteinemia
SepehrmaneshZ, KolahdoozF, AbediF, MazroiiN, AssarianA, AsemiZ, et al. (2016).
Vitamin D Supplementation Affects the Beck Depression Inventory, Insulin Resistance, and Biomarkers of Oxidative Stress in Patients with Major Depressive Disorder: A Randomized, Controlled Clinical Trial
Antidepressant activity of vorinostat is associated with amelioration of oxidative stress and inflammation in a corticosterone-induced chronic stress model in mice
Naringenin attenuates behavioral derangements induced by social defeat stress in mice via inhibition of acetylcholinesterase activity, oxidative stress and release of pro-inflammatory cytokines
ZhaoXJ, ZhaoZ, YangDD, CaoLL, ZhangL, JiJ, et al. (2017).
Activation of ATP-sensitive potassium channel by iptakalim normalizes stress-induced HPA axis disorder and depressive behaviour by alleviating inflammation and oxidative stress in mouse hypothalamus
Silymarin ameliorates experimentally induced depressive like behavior in rats: Involvement of hippocampal BDNF signaling, inflammatory cytokines and oxidative stress response
BerkiksI, BenmhammedH, MesfiouiA, OuichouA, El HasnaouiA, MoudenS, et al. (2018).
Postnatal melatonin treatment protects against affective disorders induced by early-life immune stimulation by reducing the microglia cell activation and oxidative stress
Ben SaadH, KharratN, DrissD, GargouriM, MarrakchiR, JammoussiK, et al. (2017).
Effects of vanillin on potassium bromate-induced neurotoxicity in adult mice: impact on behavior, oxidative stress, genes expression, inflammation and fatty acid composition
CasarilAM, DominguesM, BampiSR, de Andrade LourencoD, PadilhaNB, LenardaoEJ, et al. (2019).
The selenium-containing compound 3-((4-chlorophenyl)selanyl)-1-methyl-1H-indole reverses depressive-like behavior induced by acute restraint stress in mice: modulation of oxido-nitrosative stress and inflammatory pathway
Effect of 25-methoxy hispidol A isolated from Poncirus trifoliate against bacteria-induced anxiety and depression by targeting neuroinflammation, oxidative stress and apoptosis in mice
KimJS, LeeEJ, ChangDI, ParkJH, AhnSH, ChaJK, et al. (2017).
Efficacy of early administration of escitalopram on depressive and emotional symptoms and neurological dysfunction after stroke: a multicentre, double-blind, randomised, placebo-controlled study
Coming of age: molecular drivers of aging and therapeutic opportunities
1
2013
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Age and Sex Are Critical Factors in Ischemic Stroke Pathology
1
2018
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Associations between age and the course of major depressive disorder: a 2-year longitudinal cohort study
2
2018
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
... ]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association
2
2019
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
... ]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2017: a systematic analysis for the Global Burden of Disease Study 2017
1
2018
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980-2017: a systematic analysis for the Global Burden of Disease Study 2017
1
2018
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016
1
2017
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
The epidemiology of depression across cultures
1
2013
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Cross-national epidemiology of DSM-IV major depressive episode
0
2011
Depression
6
2018
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
... ]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
... In the last 5 years, mountains of preclinical and clinical studies have found that the application of antioxidants inhibits stroke and depression in patients with either, or both diseases, although the therapeutic effects and safety are under consideration. What’s more, the main pharmacotherapy for depression patients focuses on antidepressants [143], and the selective serotonin reuptake inhibitors (SSRIs) have been used as first-line antidepressants in recent decades. However, SSRIs have shown moderate efficacy and significant side-effects [10]. Variability in efficacy and acceptability are large in head-to-head trials [144]. Therefore, development of an accurate and effective antidepressant regimen requires our constant efforts. Herein, we list most of the relative studies about antioxidants and antidepressants used in treating stroke/depression, in order to find potential therapies. ...
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Reducing the global burden of depression: a Lancet-World Psychiatric Association Commission
1
2019
... Epidemiologic studies demonstrate that 11% of the world’s population is over 60 years of age and this percentage will double to about 22% by 2050 [1]. A significant percentage of older individuals develop one or more age-related diseases, which may include two leading diseases characterized by high incidence and disability: stroke [2] and depression [3]. Stroke is classified into ischemic stroke and hemorrhagic stroke; the latter consists of intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH). It is estimated that on average, a person died from stroke every 3 minutes and 42 seconds in 2016 [4]. Global Burden of Disease 2017 identifies stroke as the third leading cause of years of life lost and disability-adjusted life years [5, 6]. Between 2006 and 2016, the actual number of stroke deaths increased 3.7%, although the age-adjusted mortality rate decreased 16.7% due to the large increase in the number of elderly people [4, 7]. Like stroke, another disease that affects a significant proportion of the population is depression, a psychiatric disorder characterized by inactivity and negative feelings of inadequacy. The 12-month prevalence of major depressive disorder (MDD) is about 6%, while the lifetime risk of MDD is nearly 15-18% [8-10]. Moreover, older age is identified as a consistent and important risk factor for a worse prognosis. This phenomenon may be associated with the effect of cognitive impairment [3]. As the World Health Organization (WHO) predicted depression to be the leading cause of global burden by 2030, attention should be paid to depression treatment [10, 11]. ...
Upregulation of Nox4 by hypertrophic stimuli promotes apoptosis and mitochondrial dysfunction in cardiac myocytes
1
2010
... OS describes a state in which the body produces excessive ROS and RNS in response to deleterious substances. Under normal conditions, OS and the anti-OS system are beneficial to physiological functions, such as respiration, circulation, etc. Mitochondrial dysfunction and internal environment disturbance can arise from multiple cellular molecules and signaling pathways as a result of imbalanced redox reactions, characterized by increased production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Fig. 1), as well as decreased antioxidant defenses. The most important ROS are mitochondrial ROS (mitoROS) produced from enzymes of the mitochondrial respiratory chain. Overproduced mitoROS can affect metabolic pathways, leading to a compromised function of organelles [12]. Another significant source of ROS is NADPH oxidase (NOX). When the complex of NOX is assembled, it transports an electron from the cytosolic NADPH to oxygen to then form superoxide on the extracellular side [13]. The antioxidant system consists of enzymes and nonenzymatic substances, among which nuclear factor erythroid 2-related factor (Nrf2) is the most important factor. Nrf2 is a transcription factor and can recognize the antioxidant response element (ARE) to regulate several genes, such as heme oxygenase 1 (HO-1). Activation of the Nrf2-ARE pathway increases antioxidants and protects the cells from free radical (FR) accumulation [14]. ...
The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology
2
2007
... Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303]. ...
... OS describes a state in which the body produces excessive ROS and RNS in response to deleterious substances. Under normal conditions, OS and the anti-OS system are beneficial to physiological functions, such as respiration, circulation, etc. Mitochondrial dysfunction and internal environment disturbance can arise from multiple cellular molecules and signaling pathways as a result of imbalanced redox reactions, characterized by increased production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Fig. 1), as well as decreased antioxidant defenses. The most important ROS are mitochondrial ROS (mitoROS) produced from enzymes of the mitochondrial respiratory chain. Overproduced mitoROS can affect metabolic pathways, leading to a compromised function of organelles [12]. Another significant source of ROS is NADPH oxidase (NOX). When the complex of NOX is assembled, it transports an electron from the cytosolic NADPH to oxygen to then form superoxide on the extracellular side [13]. The antioxidant system consists of enzymes and nonenzymatic substances, among which nuclear factor erythroid 2-related factor (Nrf2) is the most important factor. Nrf2 is a transcription factor and can recognize the antioxidant response element (ARE) to regulate several genes, such as heme oxygenase 1 (HO-1). Activation of the Nrf2-ARE pathway increases antioxidants and protects the cells from free radical (FR) accumulation [14]. ...
Cerebrovascular and Neurological Disorders: Protective Role of NRF2
1
2019
... OS describes a state in which the body produces excessive ROS and RNS in response to deleterious substances. Under normal conditions, OS and the anti-OS system are beneficial to physiological functions, such as respiration, circulation, etc. Mitochondrial dysfunction and internal environment disturbance can arise from multiple cellular molecules and signaling pathways as a result of imbalanced redox reactions, characterized by increased production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Fig. 1), as well as decreased antioxidant defenses. The most important ROS are mitochondrial ROS (mitoROS) produced from enzymes of the mitochondrial respiratory chain. Overproduced mitoROS can affect metabolic pathways, leading to a compromised function of organelles [12]. Another significant source of ROS is NADPH oxidase (NOX). When the complex of NOX is assembled, it transports an electron from the cytosolic NADPH to oxygen to then form superoxide on the extracellular side [13]. The antioxidant system consists of enzymes and nonenzymatic substances, among which nuclear factor erythroid 2-related factor (Nrf2) is the most important factor. Nrf2 is a transcription factor and can recognize the antioxidant response element (ARE) to regulate several genes, such as heme oxygenase 1 (HO-1). Activation of the Nrf2-ARE pathway increases antioxidants and protects the cells from free radical (FR) accumulation [14]. ...
Oxidative & nitrosative stress in depression: why so much stress?
1
2014
... Under physiological conditions, moderate OS activity is necessary for body health. Toxic effects derived from ROS and RNS can be ameliorated or neutralized by FR scavengers and the antioxidant system. However, when a large number of ROS and RNS are generated, excessive FR then induce molecular oxidation, cell membrane modification, and enzyme inactivation, resulting in cellular damage and functional decline [15]. Overactive OS with an imbalanced redox state can induce many injuries, especially to the brain, partly expounding the role of OS in both stroke and depression. ...
Blood-brain barrier dysfunction and recovery after ischemic stroke
2
2018
... Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Mechanisms of brain injury after intracerebral haemorrhage
3
2006
... Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
... ]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Neuroprotective Role of the Nrf2 Pathway in Subarachnoid Haemorrhage and Its Therapeutic Potential
1
2019
... Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
Activation of TGR5 with INT-777 attenuates oxidative stress and neuronal apoptosis via cAMP/PKCepsilon/ALDH2 pathway after subarachnoid hemorrhage in rats
1
2019
... Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
F10, a novel hydatidiform mole-associated gene, inhibits the paclitaxel sensitivity of A549 lung cancer cells by downregulating BAX and caspase-3
1
2017
... Ischemia-hypoxia often occurs in ischemic stroke, while intracerebral bleeding in the brain parenchyma can be found in ICH, and blood clots can be found in the subarachnoid space of SAH patients. Once cerebral ischemia occurs, it impairs cellular metabolism and triggers pathological pathways, such as immune responses, inflammatory reactions, OS, autophagy, and apoptosis, leading to irreversible neuronal damage and brain injuries accompanied by blood-brain barrier (BBB) disruption (Fig. 2) [16]. Particularly, the release of FR further damages the brain. Different from ischemia, both primary and secondary brain injury (SBI) occur in ICH and SAH. Mechanisms participating in post-ICH include the coagulation cascade (particularly thrombin), hemoglobin degradation, inflammation, apoptosis, necrosis, OS, and hematoma expansion [17]. Post-ICH events lead to SBI, which manifests as brain edema, BBB disruption, brain atrophy, vasospasm, neurological deficits, and even death [17]. Compared with ischemic stroke, the results of OS activity in the studies were similar in ICH. Indeed, this kind of behavior exhibited by OS also occurs in SAH [18]. Furthermore, brain injury involved in SAH is divided into two stages: an early stage within the first 72 hours, called early brain injury (EBI), and the delayed stage. OS plays a direct and indirect role in both stages, and many recent studies demonstrate efficacy of antioxidants in animal models, especially in EBI [19, 20]. ...
HPA axis in major depression: cortisol, clinical symptomatology and genetic variation predict cognition
1
2017
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes
2
2015
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
... ], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Cortical abnormalities in adults and adolescents with major depression based on brain scans from 20 cohorts worldwide in the ENIGMA Major Depressive Disorder Working Group
1
2017
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Human Hippocampal Neurogenesis Persists throughout Aging
1
2018
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Is depression associated with increased oxidative stress? A systematic review and meta-analysis
1
2015
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Oxidative stress in major depressive and anxiety disorders, and the association with antidepressant use; results from a large adult cohort
0
2017
Oxidative stress, inflammation and treatment response in major depression
1
2017
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
The Omega-3 Index Is Inversely Associated with Depressive Symptoms among Individuals with Elevated Oxidative Stress Biomarkers
1
2016
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Serum IL-6 levels and oxidation rate of LDL cholesterol were related to depressive symptoms independent of omega-3 fatty acids among female hospital and nursing home workers in Japan
1
2019
... In comparison with stroke, the main factors promoting MDD are psychosocial stressors, and the main pathophysiology of depression is associated with decreased monoamine levels, an altered hypothalamic-pituitary-adrenal axis [21], inflammation [22], neuro-plasticity and neurogenesis controlled by BDNF [22], and structural and functional brain changes (Fig. 2) [23, 24]. Apart from inflammation and neurogenesis, OS also plays a crucial role in depression. The role of OS in depression is supported by many studies that show upregulated OS activities and downregulated anti-OS responses [25-27]. In addition, the antioxidants may ease depressive symptoms [28, 29]. In summary, there is a close relationship between depression and OS. ...
Depression after stroke at 12-month follow-up: a multicenter study
1
2017
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
Depression after minor stroke: Prevalence and predictors
2
2015
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Long-term depressive symptoms and anxiety after transient ischaemic attack or ischaemic stroke in young adults
2
2016
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Depression, anxiety, and cognitive functioning after intracerebral hemorrhage
2
2015
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Longitudinal course of depressive symptoms and anxiety after aneurysmal subarachnoid hemorrhage
2
2017
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
Depression and risk of stroke in midaged women: a prospective longitudinal study
1
2013
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
Depression and risk of stroke morbidity and mortality: a meta-analysis and systematic review
1
2011
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
Evidence of perceived psychosocial stress as a risk factor for stroke in adults: a meta-analysis
3
2015
... As mentioned above, OS not only exists in stroke, but also occurs in depression. Moreover, many clinical trials have proved that stroke increases the incidence of depression, which inversely acts as an independent risk factor for stroke. On one hand, both ischemic and hemorrhagic stroke increases the prevalence of depression. A multicenter study found that approximately one-fifth of patients developed depression after stroke at a 12-month follow-up [30]. In addition, 757 patients with first-ever minor ischemic stroke were followed for one year and analysis showed that nearly 30% of patients developed depression [31]. However, the incidence rates of depression in ischemic stroke survivors aged 18-50 years and in the control group were 16.8% and 6.1%, respectively [32]. A similar relationship is also seen in ICH [33] and SAH [34]. On the other hand, an Australian longitudinal study with a 12-year follow-up suggested that depression caused a 2-fold increase in the odds of stroke [35]. In addition, Pan did a meta-analysis consisting of 28 prospective cohort studies illustrating that the risk of ischemic stroke was exacerbated by depression [36], whereas another meta-analysis found that patients experiencing stressful life events had a 33% increased risk of total stroke [37] (Fig. 2). ...
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Tissue acidosis induces neuronal necroptosis via ASIC1a channel independent of its ionic conduction
1
2015
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Determination of Vascular Reactivity of Middle Cerebral Arteries from Stroke and Spinal Cord Injury Animal Models Using Pressure Myography
1
2016
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Significant Association Between Low Mitochondrial DNA Content in Peripheral Blood Leukocytes and Ischemic Stroke
1
2017
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Oxidative Stress Markers and Their Dynamic Changes in Patients after Acute Ischemic Stroke
1
2016
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
High Serum Levels of Malondialdehyde and 8-OHdG are both Associated with Early Cognitive Impairment in Patients with Acute Ischaemic Stroke
1
2017
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
The relationship between oxidized low-density lipoprotein and the NIHSS score among patients with acute ischemic stroke: The SOS-Stroke Study
1
2018
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Association between oxidized low-density lipoprotein and cognitive impairment in patients with ischemic stroke
1
2018
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Association of Oxidized Low-Density Lipoprotein With Prognosis of Stroke and Stroke Subtypes
1
2017
... Because the human brain requires constant oxygen and nutrients to maintain its function, it is vulnerable to FR attack. The depletion of energy in ischemic stroke can cause a series of damage to promote the development or recurrence of stroke, and OS is involved in all stages of ischemic stroke progression (Fig. 3). Firstly, energy expenditure leads to the accumulation of H+ concentration and H2O2. Studies on a mouse model of focal ischemia found that extracellular proton induce neuronal necroptosis via acid-sensing ion channel 1a/receptor interaction protein 1 association [38]. Secondly, ROS have effects on cerebral blood flow. ROS stimulates vasoconstriction and increases platelet aggregation and endothelial cell permeability, thereby affecting blood circulation [39]. Thirdly, RNS play a role in mitochondrial functions, such as reducing DNA and suppressing enzymes of the mitochondria. This is favored by a case-control study, in which a low mitochondrial DNA content in peripheral blood leukocytes is significantly related to ischemic stroke [40]. Lastly, ROS and RNS bring about DNA damage, protein destruction, lipid peroxidation, and cell death, leading to poor outcomes [41, 42]. The results of clinical studies have demonstrated that higher plasma levels of oxidized low-density lipoprotein reveal a worse prognosis [43], a higher prevalence of cognitive impairment [44], and an increased risk of death [45]. ...
Brain edema after experimental intracerebral hemorrhage: role of hemoglobin degradation products
2
2002
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
... Schematic model of oxidative mechanisms in ICH and SAH, especially associated with hemoglobin (Hb). During the hemoglobin-heme-iron axis, Hb is released into the extracellular space and is accompanied by an abundance of superoxide generated from the non-enzymatic oxidation of Hb [46]. This oxidation of Hb produces methemoglobin, which releases heme to stimulate lipid peroxidation and other oxidative actions around the hematoma in brain tissue. Meanwhile, iron released from Hb degradation is used in the Fenton reaction to transform H2O2 into the hydroxyl radical, leading to increased oxidative damage [47]. ...
Intracerebral Hematoma Contributes to Hydrocephalus After Intraventricular Hemorrhage via Aggravating Iron Accumulation
2
2015
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
... Schematic model of oxidative mechanisms in ICH and SAH, especially associated with hemoglobin (Hb). During the hemoglobin-heme-iron axis, Hb is released into the extracellular space and is accompanied by an abundance of superoxide generated from the non-enzymatic oxidation of Hb [46]. This oxidation of Hb produces methemoglobin, which releases heme to stimulate lipid peroxidation and other oxidative actions around the hematoma in brain tissue. Meanwhile, iron released from Hb degradation is used in the Fenton reaction to transform H2O2 into the hydroxyl radical, leading to increased oxidative damage [47]. ...
Efficacy of the lipid-soluble iron chelator 2,2'-dipyridyl against hemorrhagic brain injury
1
2012
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
Glial glutamate transporters expression, glutamate uptake, and oxidative stress in an experimental rat model of intracerebral hemorrhage
1
2018
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
Hemoglobin-induced oxidative stress contributes to matrix metalloproteinase activation and blood-brain barrier dysfunction in vivo
1
2010
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
Melatonin Alleviates Intracerebral Hemorrhage-Induced Secondary Brain Injury in Rats via Suppressing Apoptosis, Inflammation, Oxidative Stress, DNA Damage, and Mitochondria Injury
3
2018
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
PERK Pathway Activation Promotes Intracerebral Hemorrhage Induced Secondary Brain Injury by Inducing Neuronal Apoptosis Both in Vivo and in Vitro
1
2018
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
Distinct role of heme oxygenase-1 in early- and late-stage intracerebral hemorrhage in 12-month-old mice
1
2017
... Hemorrhagic stroke is a common, serious neurological disease associated with high disability and mortality, especially when associated with ICH. One of the main underlying mechanisms is OS. ROS and RNS usually come from the metabolite axis of hemoglobin-heme-iron, NOX activation, increased glutamate and inflammatory activities. Upon the onset of ICH, bleeding into the brain parenchyma is commonly observed. Then, several biological events associated with hemoglobin oxidation and released iron occur, resulting in neurological damage (Fig. 4) [46, 47]. What’s more, the results suggest that iron chelators could attenuate ROS generation to improve neurological function following ICH, further supporting the hemoglobin-heme-iron axis [48]. Additionally, there are other mechanisms participating in the process of ICH, such as glial glutamate transporter responses [49] and mitochondria-dependent apoptosis [50]. The effects caused by other pathological events are similar to those in ischemic stroke [51, 52]. Besides neuronal injuries, those pathological events contribute to SBI, including brain edema, BBB breakdown, and vasospasm. Interruption of the BBB facilitates ROS and RNS accumulation, which amplifies brain injuries [53]. ...
Early global brain oedema in relation to clinical admission parameters and outcome in patients with aneurysmal subarachnoid haemorrhage
1
2010
... In SAH, EBI is identified as the immediate injury accompanied by subsequent events (brain edema, inflammation, apoptosis, etc.) in the first 72 hours, and EBI plays a vital role in the pathological processes (Fig. 4). After the occurrence of SAH, the intracranial pressure will rise to the pressure of the diastolic artery, which reduces cerebrospinal fluid pressure and disturbs cerebral autoregulation before BBB breakdown and occurrence of cerebral edema [54]. Due to erythrocytosis, the central nervous system (CNS) is exposed to high levels of Hb and Hb-degradation products in the subarachnoid space. This pathological process produces excess ROS and RNS, and promotes cerebral vasospasm, cerebrovascular stenosis, and delayed cerebral ischemia [55]. Furthermore, several harmful events occur in SAH survivors, including altered ionic homeostasis, excitotoxicity, destruction of vascular integrity, OS, inflammation, apoptosis, autophagy, and activation of the NOS pathway [56-62]. ...
Definition of delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage as an outcome event in clinical trials and observational studies: proposal of a multidisciplinary research group
1
2010
... In SAH, EBI is identified as the immediate injury accompanied by subsequent events (brain edema, inflammation, apoptosis, etc.) in the first 72 hours, and EBI plays a vital role in the pathological processes (Fig. 4). After the occurrence of SAH, the intracranial pressure will rise to the pressure of the diastolic artery, which reduces cerebrospinal fluid pressure and disturbs cerebral autoregulation before BBB breakdown and occurrence of cerebral edema [54]. Due to erythrocytosis, the central nervous system (CNS) is exposed to high levels of Hb and Hb-degradation products in the subarachnoid space. This pathological process produces excess ROS and RNS, and promotes cerebral vasospasm, cerebrovascular stenosis, and delayed cerebral ischemia [55]. Furthermore, several harmful events occur in SAH survivors, including altered ionic homeostasis, excitotoxicity, destruction of vascular integrity, OS, inflammation, apoptosis, autophagy, and activation of the NOS pathway [56-62]. ...
Role of Delta133p53 isoform in NF-kappaB inhibitor PDTC-mediated growth inhibition of MKN45 gastric cancer cells
3
2017
... In SAH, EBI is identified as the immediate injury accompanied by subsequent events (brain edema, inflammation, apoptosis, etc.) in the first 72 hours, and EBI plays a vital role in the pathological processes (Fig. 4). After the occurrence of SAH, the intracranial pressure will rise to the pressure of the diastolic artery, which reduces cerebrospinal fluid pressure and disturbs cerebral autoregulation before BBB breakdown and occurrence of cerebral edema [54]. Due to erythrocytosis, the central nervous system (CNS) is exposed to high levels of Hb and Hb-degradation products in the subarachnoid space. This pathological process produces excess ROS and RNS, and promotes cerebral vasospasm, cerebrovascular stenosis, and delayed cerebral ischemia [55]. Furthermore, several harmful events occur in SAH survivors, including altered ionic homeostasis, excitotoxicity, destruction of vascular integrity, OS, inflammation, apoptosis, autophagy, and activation of the NOS pathway [56-62]. ...
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Effects of endoplasmic reticulum stress on the autophagy, apoptosis, and chemotherapy resistance of human breast cancer cells by regulating the PI3K/AKT/mTOR signaling pathway
0
2017
Baicalin Attenuates Subarachnoid Hemorrhagic Brain Injury by Modulating Blood-Brain Barrier Disruption, Inflammation, and Oxidative Damage in Mice
0
2017
Mdivi-1 ameliorates early brain injury after subarachnoid hemorrhage via the suppression of inflammation-related blood-brain barrier disruption and endoplasmic reticulum stress-based apoptosis
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Naringin alleviates early brain injury after experimental subarachnoid hemorrhage by reducing oxidative stress and inhibiting apoptosis
2
2017
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Tumor necrosis factor receptor-associated factor 6 participates in early brain injury after subarachnoid hemorrhage in rats through inhibiting autophagy and promoting oxidative stress
0
2017
The autophagy-lysosomal system in subarachnoid haemorrhage
1
2016
... In SAH, EBI is identified as the immediate injury accompanied by subsequent events (brain edema, inflammation, apoptosis, etc.) in the first 72 hours, and EBI plays a vital role in the pathological processes (Fig. 4). After the occurrence of SAH, the intracranial pressure will rise to the pressure of the diastolic artery, which reduces cerebrospinal fluid pressure and disturbs cerebral autoregulation before BBB breakdown and occurrence of cerebral edema [54]. Due to erythrocytosis, the central nervous system (CNS) is exposed to high levels of Hb and Hb-degradation products in the subarachnoid space. This pathological process produces excess ROS and RNS, and promotes cerebral vasospasm, cerebrovascular stenosis, and delayed cerebral ischemia [55]. Furthermore, several harmful events occur in SAH survivors, including altered ionic homeostasis, excitotoxicity, destruction of vascular integrity, OS, inflammation, apoptosis, autophagy, and activation of the NOS pathway [56-62]. ...
Genetic Contributions of Inflammation to Depression
1
2017
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Evidence for additionally increased apoptosis in the peripheral blood mononuclear cells of major depressive patients with a high risk for suicide
0
2018
Gut/brain axis and the microbiota
1
2015
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Depressive-Like Behaviors Are Regulated by NOX1/NADPH Oxidase by Redox Modification of NMDA Receptor 1
1
2017
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Indoleamine 2,3-Dioxygenase-Dependent Neurotoxic Kynurenine Metabolism Contributes to Poststroke Depression Induced in Mice by Ischemic Stroke along with Spatial Restraint Stress
0
2018
Nitric Oxide and Major Depressive Disorder: Pathophysiology and Treatment Implications
0
2016
Inhibition of mitochondrial respiratory chain in brain of rats subjected to an experimental model of depression
1
2008
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
A longitudinal study of neurotrophic, oxidative, and inflammatory markers in first-onset depression in midlife women
1
2018
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Markers of Oxidative Stress and Neuroprogression in Depression Disorder
1
2015
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Variation of genes involved in oxidative and nitrosative stresses in depression
1
2018
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Oxidative stress and antioxidant parameters in patients with major depressive disorder compared to healthy controls before and after antidepressant treatment: results from a meta-analysis
0
2015
Oral treatment with amitriptyline induces coenzyme Q deficiency and oxidative stress in psychiatric patients
1
2012
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Oxidative Stress and Antioxidant Parameters in Patients With Major Depressive Disorder Compared to Healthy Controls Before and After Antidepressant Treatment: Results From a Meta-Analysis
1
2015
... The impact on stroke pertains to an abundance of ROS and RNS, as well as OS-induced cell death pathways. So, what role could OS have in depression? Depression involves several pathological processes that include an imbalance of neurotransmitters, inflammation, OS, apoptosis, glutamate excitotoxicity, and the microbiota-gut-brain axis [10, 63-65]. Although there is a deficiency of direct evidence exhibiting mechanisms between OS and depression, numerous animal and human studies have reported functions of OS in depressed patients via various OS-associated molecules [66-69]. For example, Pasquali et al. conducted a longitudinal study and described a series cascade of pro-oxidative and pro-inflammatory events contributing to the development of MDD in middle-aged women [70]. Besides hyperactive OS, reduced antioxidant activity may exacerbate depression, but antioxidant treatment shows anti-depressant effects [71-74]. Wigner et al. proved that the polymorphisms of antioxidant enzymes (e.g., superoxide dismutase (SOD) and catalase (CAT) or glutathione peroxidase (GPx)) could regulate the risk of depression [72]. Parallel results in the clinical trial were found to reach a consistency in the effects of OS in depression [75]. ...
Inhibition of GSK3 differentially modulates NF-kappaB, CREB, AP-1 and beta-catenin signaling in hepatocytes, but fails to promote TNF-alpha-induced apoptosis
2
2008
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
... ]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Targeting glycogen synthase kinase-3 for oxidative stress and neuroinflammation: Opportunities, challenges and future directions for cerebral stroke management
1
2018
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
GSK-3beta downregulates Nrf2 in cultured cortical neurons and in a rat model of cerebral ischemia-reperfusion
1
2016
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Early release of HMGB-1 from neurons after the onset of brain ischemia
2
2008
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
... ]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
High-mobility group box 1 promotes metalloproteinase-9 upregulation through Toll-like receptor 4 after cerebral ischemia
1
2010
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Toll-like receptor-mediated cytokine production is differentially regulated by glycogen synthase kinase 3
1
2005
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
The immunology of stroke: from mechanisms to translation
1
2011
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Neutrophil recruitment to the brain in mouse and human ischemic stroke
1
2015
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Circulating mitochondrial DAMPs cause inflammatory responses to injury
1
2010
... Overproduction of ROS and RNS can activate inflammatory processes to aggravate brain damage through glycogen synthase kinase 3 (GSK-3) and endothelial injury. ROS stimulate the phosphorylation of GSK-3 to negatively affect the cyclic adenosine monophosphate response element-binding protein (CREB) by suppressing its nuclear translocation, resulting in an increased expression of pro-inflammatory cytokines and brain dysfunction in ischemic rat [76, 77]. To be specific, activated GSK-3 downregulates Nrf2-ARE binding activity and decreases the expression levels of genes downstream of Nrf2-ARE [78], but it stimulates the Toll-like receptors (TLRs) in the peripheral blood mononuclear cells [79]. Experiments indicate that TLR 4 induces the expression of inflammatory elements through activation and nuclear translocation of NF-κB, while deletion of TLR 4 attenuates ischemic cerebral injury [80]. Additionally, GSK-3 could boost inflammation by interrupting CREB binding proteins, which are the co-activators of CREB and NF-κB in the nucleus [76, 81]. Inversely, inhibition of GSK-3β improves the transcription and expression of anti-inflammatory cytokines and reduces pro-inflammatory cytokine production [79]. Therefore, the results of GSK-3 activation and GSK-3 suppression confirm the role of OS-induced inflammation in ischemic brain injury. In addition to GSK-3, oxidative injury of platelet and endothelium cells also participate in inflammation. Following endothelial impairment, P-selectin is highly expressed to induce the rolling of leukocytes on the endothelium, facilitating stable adhesion between leukocytes and endothelial cells [82, 83]. Afterwards, the adhered leukocytes release matrix metalloproteinase, break down the BBB, and enter into the brain parenchyma to trigger a series of inflammatory processes [84]. In ICH, a similar process occurs [51]. In SAH, free Hb stimulates endothelial cells to express cell adhesion molecules to attract neutrophils. Cells trapped in the subarachnoid space then undergo an oxidative burst, which releases ROS-mediated inflammatory cytokines or other molecules to further damage the brain [56]. ...
Inflammation in Depression and the Potential for Anti-Inflammatory Treatment
1
2016
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
Decreased Iron in Cancer Cells and Their Microenvironment Improves Cytolysis of Breast Cancer Cells by Natural Killer Cells
1
2017
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
A role for mitochondria in NLRP3 inflammasome activation
1
2011
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
NLRP3 inflammasome is activated in mononuclear blood cells from patients with major depressive disorder
1
2014
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
A neuro-immune, neuro-oxidative and neuro-nitrosative model of prenatal and postpartum depression
1
2018
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
Linking the biological underpinnings of depression: Role of mitochondria interactions with melatonin, inflammation, sirtuins, tryptophan catabolites, DNA repair and oxidative and nitrosative stress, with consequences for classification and cognition
1
2018
... In depressed individuals, ROS and RNS have effects on inflammatory reactions via NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)[85]. After sensing mitochondrial dysfunction of ROS, activated NLRP3 inflammasome induce the maturation of procaspase-1, which is an initiating process of interleukin (IL)1β and IL-18, to initiate inflammatory responses. Nevertheless, IL-1β knock-down in the hippocampus attenuate depression-like behaviors induced by LPS in mice [86]. However, these changes are reversed by treatment with amitriptyline [87, 88]. In addition, pattern recognition receptors recognize PAMPs and DAMPs to trigger MAPK and/or NF-κB to activate immune-inflammation [89, 90]. ...
Molecular mechanisms of necroptosis: an ordered cellular explosion
1
2010
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
Apoptotic and necrotic death mechanisms are concomitantly activated in the same cell after cerebral ischemia
1
2004
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
Autophagy in ischemic stroke
1
2018
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
Apoptosis-inducing factor triggered by poly(ADP-ribose) polymerase and Bid mediates neuronal cell death after oxygen-glucose deprivation and focal cerebral ischemia
2
2005
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
... ]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
miRNA-144 induces microglial autophagy and inflammation following intracerebral hemorrhage
1
2017
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
Autophagy regulates intracerebral hemorrhage induced neural damage via apoptosis and NF-kappaB pathway
1
2016
... Accumulating evidence has shown that apoptosis, necrosis, and their combined pathway, “necroptosis”, participated in ischemic stroke stimulated by ROS and RNS [91]. Experimental data obtained from ischemic mouse models suggest that the nuclei of neurons exhibit apoptotic morphology after four hours of ischemia [92]. Mechanistically, under pathological conditions, cellular stress stimulates an apoptotic signaling pathway to activate the caspase protease and cause mitochondrial dysfunction, resulting in characteristic changes of apoptotic cell morphology, such as cell rounding, plasma membrane blebbing, and nuclear fragmentation. Apoptosis can be induced by extrinsic and intrinsic stimuli. The extrinsic stimuli are triggered by the death receptor signaling pathway [93], while the intrinsic stimuli involve a mitochondrial signaling pathway and released cytochrome c (Cytc) [94]. When ischemic stroke occurs, OS initiates the apoptosis pathway mainly through intrinsic stimulation. OS also regulates apoptotic activities through modulating the balance between the anti-apoptotic protein, Bcl-2, and the pro-apoptotic regulator, Bax protein [94]. In ICH, Cytc-mediated and mitochondria-dependent apoptosis is also demonstrated to be an important part of the OS-mediated mechanisms. Additionally, Hb-induced ROS, NO, and peroxynitrite (ONOO-) are shown to directly or indirectly activate matrix metalloprotein 9, leading to apoptosis [95, 96]. Meanwhile, there is a significant increase in the number of apoptotic neural cells in the rat brain after SAH [60]. ...
"Killing the Blues": a role for cellular suicide (apoptosis) in depression and the antidepressant response?
2
2009
... Parallel to OS-mediated apoptosis in stroke, intrinsic apoptosis associated with mitochondria occupies an important position in the development of depression. When exposed to chronic stress, overactive apoptosis stimulated by OS in neurons promotes the neurodegeneration in depressed animals [97]. Mechanistically, Bcl-2-mediated damage and excess levels of Ca2+ trigger various kinases, leading to a loss of the mitochondrial membrane potential, and an increased release of intermembrane proteins into the cytoplasm. In the presence of dATP, the released Cytc binds to the cytosolic Apaf 1 to activate caspase-9 and promote apoptosome formation. Meanwhile, enhanced permeability of the outer membrane of the mitochondria allows apoptosis-inducing factor (AIF) and endonuclease G to move to the nucleus and regulate chromatinolysis [97]. In addition, chronic mild stress suppresses the expression of Bcl-2-associated athanogene-1, a gene involved in anti-apoptosis, resulting in the activation of caspases, BAX, and Bcl-2 antagonists in the mitochondria. Consequently, there is an excess of neuronal death [98]. Moreover, it is highlighted that polymorphisms of apoptotic protein genes may be associated with MDD [99]. ...
... trigger various kinases, leading to a loss of the mitochondrial membrane potential, and an increased release of intermembrane proteins into the cytoplasm. In the presence of dATP, the released Cytc binds to the cytosolic Apaf 1 to activate caspase-9 and promote apoptosome formation. Meanwhile, enhanced permeability of the outer membrane of the mitochondria allows apoptosis-inducing factor (AIF) and endonuclease G to move to the nucleus and regulate chromatinolysis [97]. In addition, chronic mild stress suppresses the expression of Bcl-2-associated athanogene-1, a gene involved in anti-apoptosis, resulting in the activation of caspases, BAX, and Bcl-2 antagonists in the mitochondria. Consequently, there is an excess of neuronal death [98]. Moreover, it is highlighted that polymorphisms of apoptotic protein genes may be associated with MDD [99]. ...
Cloning and functional analysis of BAG-1: a novel Bcl-2-binding protein with anti-cell death activity
1
1995
... Parallel to OS-mediated apoptosis in stroke, intrinsic apoptosis associated with mitochondria occupies an important position in the development of depression. When exposed to chronic stress, overactive apoptosis stimulated by OS in neurons promotes the neurodegeneration in depressed animals [97]. Mechanistically, Bcl-2-mediated damage and excess levels of Ca2+ trigger various kinases, leading to a loss of the mitochondrial membrane potential, and an increased release of intermembrane proteins into the cytoplasm. In the presence of dATP, the released Cytc binds to the cytosolic Apaf 1 to activate caspase-9 and promote apoptosome formation. Meanwhile, enhanced permeability of the outer membrane of the mitochondria allows apoptosis-inducing factor (AIF) and endonuclease G to move to the nucleus and regulate chromatinolysis [97]. In addition, chronic mild stress suppresses the expression of Bcl-2-associated athanogene-1, a gene involved in anti-apoptosis, resulting in the activation of caspases, BAX, and Bcl-2 antagonists in the mitochondria. Consequently, there is an excess of neuronal death [98]. Moreover, it is highlighted that polymorphisms of apoptotic protein genes may be associated with MDD [99]. ...
Variants in Apaf-1 segregating with major depression promote apoptosome function
1
2006
... Parallel to OS-mediated apoptosis in stroke, intrinsic apoptosis associated with mitochondria occupies an important position in the development of depression. When exposed to chronic stress, overactive apoptosis stimulated by OS in neurons promotes the neurodegeneration in depressed animals [97]. Mechanistically, Bcl-2-mediated damage and excess levels of Ca2+ trigger various kinases, leading to a loss of the mitochondrial membrane potential, and an increased release of intermembrane proteins into the cytoplasm. In the presence of dATP, the released Cytc binds to the cytosolic Apaf 1 to activate caspase-9 and promote apoptosome formation. Meanwhile, enhanced permeability of the outer membrane of the mitochondria allows apoptosis-inducing factor (AIF) and endonuclease G to move to the nucleus and regulate chromatinolysis [97]. In addition, chronic mild stress suppresses the expression of Bcl-2-associated athanogene-1, a gene involved in anti-apoptosis, resulting in the activation of caspases, BAX, and Bcl-2 antagonists in the mitochondria. Consequently, there is an excess of neuronal death [98]. Moreover, it is highlighted that polymorphisms of apoptotic protein genes may be associated with MDD [99]. ...
Oxidative Stress and the Microbiota-Gut-Brain Axis. Oxidative Medicine and
1
2018
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis
1
2018
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
Dysphagia after stroke: incidence, diagnosis, and pulmonary complications
1
2005
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
New-onset fecal incontinence after stroke: prevalence, natural history, risk factors, and impact
1
2003
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
Gastrointestinal bleeding after acute ischemic stroke
1
2008
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
New-Onset Constipation at Acute Stage After First Stroke Incidence, Risk Factors, and Impact on the Stroke Outcome
1
2009
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
An unexplored brain-gut microbiota axis in stroke
1
2017
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
Commensal microbiota affects ischemic stroke outcome by regulating intestinal gammadelta T cells
1
2016
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
Principles and clinical implications of the brain-gut-enteric microbiota axis
1
2009
... Recently, the microbiota-gut-brain axis has gained extensive attention as a channel for communication and physiological regulation. Activities of the intestinal microbiome might promote abnormal protein aggregation and oxidative responses to impair the brain. Inversely, the brain can either directly or indirectly impact commensal organisms or gastrointestinal functions through the release of signaling molecules from cells in the lamina propria [100, 101]. After an ischemic stroke, there is an increase of gastrointestinal complications such as dysphagia [102], fecal incontinence [103], gastro-intestinal hemorrhage [104], and constipation [105]. Wen et al. revealed that ischemic stroke exacerbates gut barrier breakdown and microbiota alterations. Subsequently, the translocation of a selective bacterial strain from the host intestinal microbiota to the surrounding tissues promotes post-stroke infections [106]. Furthermore, Benakis C et al. suggested that intestinal dysbiosis disequilibrates the intestinal immune homeostasis by inhibiting interleukin-17-positive γδ T cells, and suppressing the trafficking of effector T cells from the gut to the leptomeninges [107]. Moreover, vagal afferent innervation of the enterochromaffin cells provides direct signaling to the neuronal circuits, leading to changes of the immune-response and other homeostatic functions [108]. Due to lack of data showing the relationship the among gut-brain axis and ICH or SAH, further studies are needed. ...
Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host's metabolism
1
2016
... In depression, the changes of microbiota, intestinal composition, and the resultant bacterial translocation expand the content of two-way interaction of the gut-brain axis. Zhang P et al. recognized the gut microbiome as an environmental factor that can shape the brain through this axis. Data from clinical sampling showed a significant difference of gut microbiota compositions between MDD patients and healthy controls, demonstrating that gut dysbiosis may mediate the host's metabolism to promote depressive-like behaviors [109]. In addition, gut dysbiosis could have an impact on the biology of MDD through several pathways involving immune activation, ROS and RNS, and neuroplasticity cascades [110]. Therefore, the microbiota-gut-brain axis may play a key role in the pathophysiological process of MDD. This axis opens a new preventative and therapeutic perspective for depression. However, due to methodological inconsistencies and limitations, more fundamental and clinical research should be undertaken. ...
Gut Microbiota, Bacterial Translocation, and Interactions with Diet: Pathophysiological Links between Major Depressive Disorder and Non-Communicable Medical Comorbidities
1
2017
... In depression, the changes of microbiota, intestinal composition, and the resultant bacterial translocation expand the content of two-way interaction of the gut-brain axis. Zhang P et al. recognized the gut microbiome as an environmental factor that can shape the brain through this axis. Data from clinical sampling showed a significant difference of gut microbiota compositions between MDD patients and healthy controls, demonstrating that gut dysbiosis may mediate the host's metabolism to promote depressive-like behaviors [109]. In addition, gut dysbiosis could have an impact on the biology of MDD through several pathways involving immune activation, ROS and RNS, and neuroplasticity cascades [110]. Therefore, the microbiota-gut-brain axis may play a key role in the pathophysiological process of MDD. This axis opens a new preventative and therapeutic perspective for depression. However, due to methodological inconsistencies and limitations, more fundamental and clinical research should be undertaken. ...
Physical inactivity is a strong risk factor for stroke in the oldest old: Findings from a multi-ethnic population (the Northern Manhattan Study)
1
2017
... The preceding content shows us a close connection between stroke and depression via the OS system and OS-mediated biological processes. Moreover, numerous studies have proved that aging or senescence is a risk factor that aggravates stroke and depression. For example, the population-based multiethnic cohort reported that the risk effect of physical inactivity is modified by age, and there is a conspicuous risk found only in stroke patients >80 years of age [111, 112]. Population-based surveillance studies predicted that the number of strokes will double by 2050, with incidence occurring primarily in the elderly (>age 75) groups [113]. Although depression prevalence in the elderly varies across studies, Sjoberg L et al. indicated that older adults with poor physical function have a higher prevalence of depression [114]. Herein, there is an “aging-stroke-depression” network and OS seems to be the center of this network. So, what relationship does aging have with oxidative stress, stroke, and depression? ...
Pathogenic ischemic stroke phenotypes in the NINDS-stroke genetics network
1
2014
... The preceding content shows us a close connection between stroke and depression via the OS system and OS-mediated biological processes. Moreover, numerous studies have proved that aging or senescence is a risk factor that aggravates stroke and depression. For example, the population-based multiethnic cohort reported that the risk effect of physical inactivity is modified by age, and there is a conspicuous risk found only in stroke patients >80 years of age [111, 112]. Population-based surveillance studies predicted that the number of strokes will double by 2050, with incidence occurring primarily in the elderly (>age 75) groups [113]. Although depression prevalence in the elderly varies across studies, Sjoberg L et al. indicated that older adults with poor physical function have a higher prevalence of depression [114]. Herein, there is an “aging-stroke-depression” network and OS seems to be the center of this network. So, what relationship does aging have with oxidative stress, stroke, and depression? ...
Population shifts and the future of stroke: forecasts of the future burden of stroke
1
2012
... The preceding content shows us a close connection between stroke and depression via the OS system and OS-mediated biological processes. Moreover, numerous studies have proved that aging or senescence is a risk factor that aggravates stroke and depression. For example, the population-based multiethnic cohort reported that the risk effect of physical inactivity is modified by age, and there is a conspicuous risk found only in stroke patients >80 years of age [111, 112]. Population-based surveillance studies predicted that the number of strokes will double by 2050, with incidence occurring primarily in the elderly (>age 75) groups [113]. Although depression prevalence in the elderly varies across studies, Sjoberg L et al. indicated that older adults with poor physical function have a higher prevalence of depression [114]. Herein, there is an “aging-stroke-depression” network and OS seems to be the center of this network. So, what relationship does aging have with oxidative stress, stroke, and depression? ...
Prevalence of depression: Comparisons of different depression definitions in population-based samples of older adults
1
2017
... The preceding content shows us a close connection between stroke and depression via the OS system and OS-mediated biological processes. Moreover, numerous studies have proved that aging or senescence is a risk factor that aggravates stroke and depression. For example, the population-based multiethnic cohort reported that the risk effect of physical inactivity is modified by age, and there is a conspicuous risk found only in stroke patients >80 years of age [111, 112]. Population-based surveillance studies predicted that the number of strokes will double by 2050, with incidence occurring primarily in the elderly (>age 75) groups [113]. Although depression prevalence in the elderly varies across studies, Sjoberg L et al. indicated that older adults with poor physical function have a higher prevalence of depression [114]. Herein, there is an “aging-stroke-depression” network and OS seems to be the center of this network. So, what relationship does aging have with oxidative stress, stroke, and depression? ...
Oxidants, antioxidants, and the degenerative diseases of aging
1
1993
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Oxidative stress in brain aging. Implications for therapeutics of neurodegenerative diseases
1
2002
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
The effects of aging in the hippocampus and cognitive decline
1
2017
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Nutrients, Microglia Aging, and Brain Aging
1
2016
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Nutritional Factors Affecting Adult Neurogenesis and Cognitive Function
1
2017
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Reduced Nrf2 expression mediates the decline in neural stem cell function during a critical middle-age period
1
2016
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Potentiation of glutathione loss and nerve cell death by the transition metals iron and copper: Implications for age-related neurodegenerative diseases
1
2018
... There is a close link between oxidative stress and aging, and this link can be proven through many related mechanisms [115, 116]. Firstly, age-related cognitive decline is a consequence of increased OS and neuroinflammation activity in the the aging hippocampus, and a consequence of reduced neurogenesis and synaptic plasticity [117]. Furthermore, mutual effects of inflammation and OS are observed to exacerbate the aging brain. Inflammation stimulates both macrophages and microglia to generate mitoROS to cause cognitive decline, whereas OS-damaged cells produce inflammatory mediators to promote microglial aging [118]. Secondly, aging and OS can damage the brain by negatively affecting neuroplasticity, brain homeostasis, and cognitive function [119, 120]. The third factor is glutathione. In animal experiments, glutathione deficiency might compromise the ability of the aging brain to meet the demands of OS, leading to impaired physiological functions [121]. ...
Hallmarks of Brain Aging: Adaptive and Pathological Modification by Metabolic States
2
2018
... Normally, physiological events of aging involve a decline of innate functions, including shortened telomeres, dysregulated hormones, and dampened immune responses. Under pathological conditions, aging stimulates and exacerbates cellular injuries, such as DNA damage and FR accumulation, to increase the vulnerability of the brain [122]. Mattson MP et al. depicted ten hallmarks of brain aging (e.g., OS and impaired DNA repair), among which, dysregulated energy metabolism is the core factor [122]. When exposed to ischemia, aging impairs the integrity of the neurovascular unit and damages brain tissues [123]. Aging can also destroy collateral circulation and revascularization of the brain through increased FR and inflammatory responses to aggravate stroke [124]. Age-related cognitive deteriorations also exist in the ICH mice model. Researchers showed a delayed neurological improvement and decreased levels of antioxidants in senescence-accelerated mice than senescence-accelerated resistant mice, though the time period in which the neurological deficits occurred and increased remains the same. To conclude, these results implied a key role of OS in senescence and stroke. ...
... ]. Mattson MP et al. depicted ten hallmarks of brain aging (e.g., OS and impaired DNA repair), among which, dysregulated energy metabolism is the core factor [122]. When exposed to ischemia, aging impairs the integrity of the neurovascular unit and damages brain tissues [123]. Aging can also destroy collateral circulation and revascularization of the brain through increased FR and inflammatory responses to aggravate stroke [124]. Age-related cognitive deteriorations also exist in the ICH mice model. Researchers showed a delayed neurological improvement and decreased levels of antioxidants in senescence-accelerated mice than senescence-accelerated resistant mice, though the time period in which the neurological deficits occurred and increased remains the same. To conclude, these results implied a key role of OS in senescence and stroke. ...
Mitochondrial function in hypoxic ischemic injury and influence of aging
2
2017
... Normally, physiological events of aging involve a decline of innate functions, including shortened telomeres, dysregulated hormones, and dampened immune responses. Under pathological conditions, aging stimulates and exacerbates cellular injuries, such as DNA damage and FR accumulation, to increase the vulnerability of the brain [122]. Mattson MP et al. depicted ten hallmarks of brain aging (e.g., OS and impaired DNA repair), among which, dysregulated energy metabolism is the core factor [122]. When exposed to ischemia, aging impairs the integrity of the neurovascular unit and damages brain tissues [123]. Aging can also destroy collateral circulation and revascularization of the brain through increased FR and inflammatory responses to aggravate stroke [124]. Age-related cognitive deteriorations also exist in the ICH mice model. Researchers showed a delayed neurological improvement and decreased levels of antioxidants in senescence-accelerated mice than senescence-accelerated resistant mice, though the time period in which the neurological deficits occurred and increased remains the same. To conclude, these results implied a key role of OS in senescence and stroke. ...
... There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders? ...
Aerobic exercise prevents rarefaction of pial collaterals and increased stroke severity that occur with aging
1
2017
... Normally, physiological events of aging involve a decline of innate functions, including shortened telomeres, dysregulated hormones, and dampened immune responses. Under pathological conditions, aging stimulates and exacerbates cellular injuries, such as DNA damage and FR accumulation, to increase the vulnerability of the brain [122]. Mattson MP et al. depicted ten hallmarks of brain aging (e.g., OS and impaired DNA repair), among which, dysregulated energy metabolism is the core factor [122]. When exposed to ischemia, aging impairs the integrity of the neurovascular unit and damages brain tissues [123]. Aging can also destroy collateral circulation and revascularization of the brain through increased FR and inflammatory responses to aggravate stroke [124]. Age-related cognitive deteriorations also exist in the ICH mice model. Researchers showed a delayed neurological improvement and decreased levels of antioxidants in senescence-accelerated mice than senescence-accelerated resistant mice, though the time period in which the neurological deficits occurred and increased remains the same. To conclude, these results implied a key role of OS in senescence and stroke. ...
Life and death in the trash heap: The ubiquitin proteasome pathway and UCHL1 in brain aging, neurodegenerative disease and cerebral Ischemia
1
2017
... Apart from OS, several other mechanisms are involved in OS and stroke. The ubiquitin proteasome pathway protects neurons by removing abnormal or toxic proteins located in the axons and dendrites, and ischemia may exacerbate injury of an aged brain through impairing ubiquitin proteasome function [125]. Furthermore, aging is detrimental to ischemic stroke recovery due to its role in altering astrocytic proliferation, inhibiting vascular endothelial growth factor (VEGF) production, and upregulating the release of inflammatory cytokines [126, 127]. What about aging in hemorrhagic stroke? Apart from the direct effects of aging in ICH, senescence may contribute to ICH progression by changing the body physiology and increasing the risk of multiple chronic health conditions and comorbidities, such as hypertension and diabetes [128]. However, the exact mechanism remains unclear. To our knowledge, there is no clear evidence explaining a relation between SAH and aging. ...
Accelerated glial reactivity to stroke in aged rats correlates with reduced functional recovery
1
2003
... Apart from OS, several other mechanisms are involved in OS and stroke. The ubiquitin proteasome pathway protects neurons by removing abnormal or toxic proteins located in the axons and dendrites, and ischemia may exacerbate injury of an aged brain through impairing ubiquitin proteasome function [125]. Furthermore, aging is detrimental to ischemic stroke recovery due to its role in altering astrocytic proliferation, inhibiting vascular endothelial growth factor (VEGF) production, and upregulating the release of inflammatory cytokines [126, 127]. What about aging in hemorrhagic stroke? Apart from the direct effects of aging in ICH, senescence may contribute to ICH progression by changing the body physiology and increasing the risk of multiple chronic health conditions and comorbidities, such as hypertension and diabetes [128]. However, the exact mechanism remains unclear. To our knowledge, there is no clear evidence explaining a relation between SAH and aging. ...
Increased levels of proinflammatory cytokines in the aged rat brain attenuate injury-induced cytokine response after excitotoxic damage
1
2009
... Apart from OS, several other mechanisms are involved in OS and stroke. The ubiquitin proteasome pathway protects neurons by removing abnormal or toxic proteins located in the axons and dendrites, and ischemia may exacerbate injury of an aged brain through impairing ubiquitin proteasome function [125]. Furthermore, aging is detrimental to ischemic stroke recovery due to its role in altering astrocytic proliferation, inhibiting vascular endothelial growth factor (VEGF) production, and upregulating the release of inflammatory cytokines [126, 127]. What about aging in hemorrhagic stroke? Apart from the direct effects of aging in ICH, senescence may contribute to ICH progression by changing the body physiology and increasing the risk of multiple chronic health conditions and comorbidities, such as hypertension and diabetes [128]. However, the exact mechanism remains unclear. To our knowledge, there is no clear evidence explaining a relation between SAH and aging. ...
The role of age in intracerebral hemorrhages
1
2015
... Apart from OS, several other mechanisms are involved in OS and stroke. The ubiquitin proteasome pathway protects neurons by removing abnormal or toxic proteins located in the axons and dendrites, and ischemia may exacerbate injury of an aged brain through impairing ubiquitin proteasome function [125]. Furthermore, aging is detrimental to ischemic stroke recovery due to its role in altering astrocytic proliferation, inhibiting vascular endothelial growth factor (VEGF) production, and upregulating the release of inflammatory cytokines [126, 127]. What about aging in hemorrhagic stroke? Apart from the direct effects of aging in ICH, senescence may contribute to ICH progression by changing the body physiology and increasing the risk of multiple chronic health conditions and comorbidities, such as hypertension and diabetes [128]. However, the exact mechanism remains unclear. To our knowledge, there is no clear evidence explaining a relation between SAH and aging. ...
The Depressed Frail Phenotype: The Clinical Manifestation of Increased Biological Aging
1
2016
... In the process of aging, biological changes in depression include mitochondrial dysfunction, dopamine dysfunction, and increased proinflammatory cytokines involved in the cellular senescence cycle. In general, mitochondrial abnormalities can be measured in peripheral blood mononuclear cells [129]. For instance, the clinical study by Karabatsiakis et al. depicted higher levels of mitochondrial impairments in older folks with depression when compared with healthy controls [130]. A systematic review and meta-analysis also supported this connection, indicating that late-life depression may be associated with a decreased hippocampal volume, though the relationship was not straightforward [131]. ...
Mitochondrial respiration in peripheral blood mononuclear cells correlates with depressive subsymptoms and severity of major depression
1
2014
... In the process of aging, biological changes in depression include mitochondrial dysfunction, dopamine dysfunction, and increased proinflammatory cytokines involved in the cellular senescence cycle. In general, mitochondrial abnormalities can be measured in peripheral blood mononuclear cells [129]. For instance, the clinical study by Karabatsiakis et al. depicted higher levels of mitochondrial impairments in older folks with depression when compared with healthy controls [130]. A systematic review and meta-analysis also supported this connection, indicating that late-life depression may be associated with a decreased hippocampal volume, though the relationship was not straightforward [131]. ...
Late-Life Depression, Hippocampal Volumes, and Hypothalamic-Pituitary-Adrenal Axis Regulation: A Systematic Review and Meta-analysis
1
2017
... In the process of aging, biological changes in depression include mitochondrial dysfunction, dopamine dysfunction, and increased proinflammatory cytokines involved in the cellular senescence cycle. In general, mitochondrial abnormalities can be measured in peripheral blood mononuclear cells [129]. For instance, the clinical study by Karabatsiakis et al. depicted higher levels of mitochondrial impairments in older folks with depression when compared with healthy controls [130]. A systematic review and meta-analysis also supported this connection, indicating that late-life depression may be associated with a decreased hippocampal volume, though the relationship was not straightforward [131]. ...
Association between decline in brain dopamine activity with age and cognitive and motor impairment in healthy individuals
1
1998
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Dopamine transporter PET in normal aging: dopamine transporter decline and its possible role in preservation of motor function
1
2010
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Striatal dopamine transporters correlate with simple reaction time in elderly subjects
1
2008
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Relationship of interleukin-6 and tumor necrosis factor-alpha with muscle mass and muscle strength in elderly men and women: the Health ABC Study
1
2002
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Subjective wellbeing, health, and ageing
2
2015
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
... ]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Depression, malnutrition, and health-related quality of life among Nepali older patients
1
2018
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
Folates and aging: Role in mild cognitive impairment, dementia and depression
1
2015
... Different from other mechanisms in stroke, the pathological process of depression during aging is significantly affected by dopamine dysfunction, inflammation, and psychological stressors. In regards to dopamine, the availability of D2 receptors in the caudate and shell nuclei is attenuated in older adults, leading to decreased motor speed and impaired frontal functioning [132]. Worse still, dopamine levels in the striatum of the elderly are only 40% of those found in young adults, and the D1/D2 receptor density and dopamine transporter expression decreased approximately 10% per decade, throughout the life cycle [133, 134]. Secondly, studies have found that inflammatory processes, especially high expression levels of IL-6, TNF-α, and C-reactive protein contribute to the depressive symptoms in elderly adults, with clinical manifestations of lethargy, slowing, and weakness [135]. Most importantly, psychological well-being is largely associated with the onset and progression of depression among the elderly population [136]. As the relationship between physical health and subjective well-being is bidirectional, elderly people with chronic diseases (e.g, coronary heart disease and chronic lung disease) have higher levels of unhappiness and are more likely to have depression [136]. Fourthly, a cross-sectional survey showed that malnutrition increased the prevalence of depression in the elderly Nepali population. [137]. Lastly, low folate levels result in increased mild cognitive impairment, dementia, and depression in older individuals [138]. In summary, aging and depression are closely linked through OS, OS-related inflammation, and dopamine dysfunction, although social life and malnutrition play a crucial role in this link. ...
The NAD+/PARP1/SIRT1 Axis in Aging
1
2017
... There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders? ...
E2F transcription factor 1 regulates cellular and organismal senescence by inhibiting Forkhead box O transcription factors
1
2014
... There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders? ...
Neuroprotective effects of the catalytic subunit of telomerase: A potential therapeutic target in the central nervous system
1
2016
... There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders? ...
Oxidative stress and depressive symptoms in older adults: A magnetic resonance spectroscopy study
1
2015
... There are some changes of molecules and signaling pathways in aging. Under ischemia/hypoxia conditions among elderly individuals, silent information regulator 1 (SIRT1) expression and the mitochondrial unfolded protein response are reduced with age, leading to impairment of mitochondrial function [139]. This condition also occurs in another study, indicating that the post-translational regulation of molecular mediators, such as hypoxia-inducible factor 1α and SIRT1, and the glycolytic-mitochondrial energy axis are critical in response to hypoxic-ischemic injury [123]. Moreover, E2f transcription factor 1 enhances the cellular senescence in human fibroblast cells, while transcription factor FOXO3 plays against senescence by regulating ROS scavenging proteins. E2f transcription factor 1 could inhibit FOXO3-dependent transcription by directly binding to FOXO3 in the nucleus to expedite the aging process [140]. Interestingly, the telomerase reverse transcriptase (TERT), a catalytic subunit of telomerase, exerts neuroprotective effects in the mitochondria of neurons by decreasing the ROS and protecting DNA. Nevertheless, recent studies observe the neuroprotective effects of TERT in cellular and animal models after aged brain injury, but it is unclear about TERT’s effects in the human CNS [141]. In regards to aging, OS, and depression, glutathione captures our attention. In a study enrolling 58 depressed older patients and 12 controls, glutathione was measured in the anterior cingulate cortex, and the data showed that increased glutathione/creatine ratios are associated with greater depressive symptoms than the control group [142]. In conclusion, OS lies in the center of the “aging-stroke-depression” network (Fig. 6). First, when stroke occurs in animals or patients, excessive generation of ROS follows, leading to cellular damage and brain injury. Second, OS mediates inflammation, apoptosis, and the microbiota-gut-brain axis to increase the accumulation of ROS, followed by brain deterioration. Third, aging acts as a risk factor and aggravates the development of stroke and depression via OS and OS-induced pathways. Due to the central role of OS in this network, administration of antioxidants seems to provide therapeutic ways for stroke and depression. So, are antioxidants safe and effective in these two disorders? ...
Management of Depression in Older Adults: A Review
1
2017
... In the last 5 years, mountains of preclinical and clinical studies have found that the application of antioxidants inhibits stroke and depression in patients with either, or both diseases, although the therapeutic effects and safety are under consideration. What’s more, the main pharmacotherapy for depression patients focuses on antidepressants [143], and the selective serotonin reuptake inhibitors (SSRIs) have been used as first-line antidepressants in recent decades. However, SSRIs have shown moderate efficacy and significant side-effects [10]. Variability in efficacy and acceptability are large in head-to-head trials [144]. Therefore, development of an accurate and effective antidepressant regimen requires our constant efforts. Herein, we list most of the relative studies about antioxidants and antidepressants used in treating stroke/depression, in order to find potential therapies. ...
Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: a systematic review and network meta-analysis
1
2018
... In the last 5 years, mountains of preclinical and clinical studies have found that the application of antioxidants inhibits stroke and depression in patients with either, or both diseases, although the therapeutic effects and safety are under consideration. What’s more, the main pharmacotherapy for depression patients focuses on antidepressants [143], and the selective serotonin reuptake inhibitors (SSRIs) have been used as first-line antidepressants in recent decades. However, SSRIs have shown moderate efficacy and significant side-effects [10]. Variability in efficacy and acceptability are large in head-to-head trials [144]. Therefore, development of an accurate and effective antidepressant regimen requires our constant efforts. Herein, we list most of the relative studies about antioxidants and antidepressants used in treating stroke/depression, in order to find potential therapies. ...
Neuroprotective role of fucoxanthin against cerebral ischemic/reperfusion injury through activation of Nrf2/HO-1 signaling
1
2018
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Sirtuin 6 protects the brain from cerebral ischemia/reperfusion injury through NRF2 activation
1
2017
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Protocatechualdehyde Protects Against Cerebral Ischemia-Reperfusion-Induced Oxidative Injury Via Protein Kinase Cepsilon/Nrf2/HO-1 Pathway
1
2017
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Protective effects of mangiferin on cerebral ischemia-reperfusion injury and its mechanisms
1
2016
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Pretreatment with Korean red ginseng or dimethyl fumarate attenuates reactive gliosis and confers sustained neuroprotection against cerebral hypoxic-ischemic damage by an Nrf2-dependent mechanism
1
2019
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Posttreatment with 11-Keto-beta-Boswellic Acid Ameliorates Cerebral Ischemia-Reperfusion Injury: Nrf2/HO-1 Pathway as a Potential Mechanism
1
2015
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Pre-treatment with metformin activates Nrf2 antioxidant pathways and inhibits inflammatory responses through induction of AMPK after transient global cerebral ischemia
1
2015
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
S-allyl cysteine activates the Nrf2-dependent antioxidant response and protects neurons against ischemic injury in vitro and in vivo
1
2015
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Neuro-protective effect of monomethyl fumarate on ischemia reperfusion injury in rats: Role of Nrf2/HO1 pathway in peri-infarct region
1
2019
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Neuroprotective mechanisms of dieckol against glutamate toxicity through reactive oxygen species scavenging and nuclear factor-like 2/heme oxygenase-1 pathway
1
2019
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Fumarate modulates the immune/inflammatory response and rescues nerve cells and neurological function after stroke in rats
1
2016
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Uric Acid Protects against Focal Cerebral Ischemia/Reperfusion-Induced Oxidative Stress via Activating Nrf2 and Regulating Neurotrophic Factor Expression
1
2018
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
A Dual AMPK/Nrf2 Activator Reduces Brain Inflammation After Stroke by Enhancing Microglia M2 Polarization
1
2018
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Andrographolide stimulates p38 mitogen-activated protein kinase-nuclear factor erythroid-2-related factor 2-heme oxygenase 1 signaling in primary cerebral endothelial cells for definite protection against ischemic stroke in rats
1
2016
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Nitroxide antioxidant as a potential strategy to attenuate the oxidative/nitrosative stress induced by hydrogen peroxide plus nitric oxide in cultured neurons. Nitric Oxide-Biology and
1
2016
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
3H-1,2-Dithiole-3-thione as a novel therapeutic agent for the treatment of ischemic stroke through Nrf2 defense pathway
1
2017
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
3-n-butylphthalide exerts neuroprotective effects by enhancing anti-oxidation and attenuating mitochondrial dysfunction in an in vitro model of ischemic stroke
1
2018
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Post-ischemic administration of 5-methoxyindole-2-carboxylic acid at the onset of reperfusion affords neuroprotection against stroke injury by preserving mitochondrial function and attenuating oxidative stress
1
2018
... Antioxidants or anti-oxidative systems have neuroprotective effects through different anti-activities and various signaling pathways, especially the Nrf2 pathway found in either ischemic stroke or hemorrhagic stroke (Table 1). Firstly, antioxidants alleviate ischemic stroke by attenuating the OS activity, and by protecting the mitochondrial function and BBB. For example, fucoxanthin [145], Sirtuin 6 [146], protocatechualdehyde [147], mangiferin [148], Korean Red Ginseng [149], 11-Keto-β-boswellic acid [150], metformin [151], s-allyl cysteine [152], monomethyl fumarate[153], dieckol [154], and fumarate [155] are reported to protect the brain by activating the Nrf2/HO-1 pathway. As such, uric acid (UA) [156], HP-1c [157], andrographolide [158], 2,2,6,6-tetramethyl-1-piperidinoxyl [159], and 3H-1,2-Dithiole-3-thione are associated with Nrf2 [160]. Secondly, protecting mitochondrial function is beneficial to ischemic stroke treatment, such as 3-n-butylphthalide [161] and 5-methoxyindole-2-carboxylic acid [162]. ...
Melatonin prevents cell death and mitochondrial dysfunction via a SIRT1-dependent mechanism during ischemic-stroke in mice
1
2015
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Progesterone induces neuroprotection following reperfusion-promoted mitochondrial dysfunction after focal cerebral ischemia in rats
1
2017
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Neuroprotective Effects of Mitochondria-Targeted Plastoquinone in a Rat Model of Neonatal Hypoxic(-)Ischemic Brain Injury
1
2018
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Ginkgolide K promotes angiogenesis in a middle cerebral artery occlusion mouse model via activating JAK2/STAT3 pathway
1
2018
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Protective effects of leonurine against ischemic stroke in mice by activating nuclear factor erythroid 2-related factor 2 pathway
1
2019
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Astragaloside IV protects blood-brain barrier integrity from LPS-induced disruption via activating Nrf2 antioxidant signaling pathway in mice
1
2018
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Osthole, a natural coumarin improves cognitive impairments and BBB dysfunction after transient global brain ischemia in C57 BL/6J mice: involvement of Nrf2 pathway
1
2015
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Panax notoginseng Saponins Protect Cerebral Microvascular Endothelial Cells against Oxygen-Glucose Deprivation/Reperfusion-Induced Barrier Dysfunction via Activation of PI3K/Akt/Nrf2 Antioxidant Signaling Pathway
1
2018
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Chinese herbal medicine formula tao hong si wu decoction protects against cerebral ischemia-reperfusion injury via PI3K/Akt and the Nrf2 signaling pathway
1
2015
... Melatonin activates the silent information regulator 1 signaling [163], and progesterone suppresses mtROS production and blocks MPTP, to exert mitochondrial protection [164]. SkQR1 is a mitochondrial-targeted antioxidant, and it is effective in protecting newborn brains before and after ischemic stroke [165]. Thirdly, antioxidant systems also exert protective effects on blood vessels. For instance, ginkgolide [166] and leonurine [167] promote angiogenesis via upregulated VEGF expression. Fourthly, BBB integrity can be protected by several antioxidants, including astragaloside IV [168], Osthole [169] and the Chinese herbal medicine formula, Tao Hong Si Wu Decoction[170, 171]. All of these antioxidants are correlated with Nrf2 signaling. ...
Schizandrin A protects against cerebral ischemia-reperfusion injury by suppressing inflammation and oxidative stress and regulating the AMPK/Nrf2 pathway regulation
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Tryptanthrin Suppresses the Activation of the LPS-Treated BV2 Microglial Cell Line via Nrf2/HO-1 Antioxidant Signaling
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Neuroprotective effects of andrographolide derivative CX-10 in transient focal ischemia in rat: Involvement of Nrf2/AE and TLR/NF-kappaB signaling
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Quercetin inhibits LPS-induced adhesion molecule expression and oxidant production in human aortic endothelial cells by p38-mediated Nrf2 activation and antioxidant enzyme induction
1
2016
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Dioscin inhibits ischemic stroke-induced inflammation through inhibition of the TLR4/MyD88/NF-B signaling pathway in a rat model
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Nrf2 inhibits NLRP3 inflammasome activation through regulating Trx1/TXNIP complex in cerebral ischemia reperfusion injury
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
The Zinc Ion Chelating Agent TPEN Attenuates Neuronal Death/apoptosis Caused by Hypoxia/ischemia Via Mediating the Pathophysiological Cascade Including Excitotoxicity, Oxidative Stress, and Inflammation
1
2015
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Inhibition of Myeloperoxidase by N-Acetyl Lysyltyrosylcysteine Amide Reduces Oxidative Stress-Mediated Inflammation, Neuronal Damage, and Neural Stem Cell Injury in a Murine Model of Stroke
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Tanshinone IIA Elicits Neuroprotective Effect Through Activating the Nuclear Factor Erythroid 2-Related Factor-Dependent Antioxidant Response
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Neuroprotection of Resveratrol Against Focal Cerebral Ischemia/Reperfusion Injury in Mice Through a Mechanism Targeting Gut-Brain Axis
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
EPO-cyclosporine combination therapy reduced brain infarct area in rat after acute ischemic stroke: role of innate immune-inflammatory response, micro-RNAs and MAPK family signaling pathway
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Rhein protects against cerebral ischemic/reperfusioninduced oxidative stress and apoptosis in rats
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Cancer stemness and metastatic potential of the novel tumor cell line K3: an inner mutated cell of bone marrow-derived mesenchymal stem cells
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Acteoside Attenuates Oxidative Stress and Neuronal Apoptosis in Rats with Focal Cerebral Ischemia-Reperfusion Injury
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Pomalidomide Ameliorates H(2)O(2)-Induced Oxidative Stress Injury and Cell Death in Rat Primary Cortical Neuronal Cultures by Inducing Anti-Oxidative and Anti-Apoptosis Effects
2
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
... ] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis
1
2016
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Adiponectin Attenuates Oxygen-Glucose Deprivation-Induced Mitochondrial Oxidative Injury and Apoptosis in Hippocampal HT22 Cells via the JAK2/STAT3 Pathway
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
YiQiFuMai Powder Injection Protects against Ischemic Stroke via Inhibiting Neuronal Apoptosis and PKCdelta/Drp1-Mediated Excessive Mitochondrial Fission
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Epigallocatechin-3-Gallate Reduces Neuronal Apoptosis in Rats after Middle Cerebral Artery Occlusion Injury via PI3K/AKT/eNOS Signaling Pathway
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Cocktail of Four Active Components Derived from Sheng Mai San Inhibits Hydrogen Peroxide-Induced PC12 Cell Apoptosis Linked with the Caspase-3/ROCK1/MLC Pathway
1
2015
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Diallyl trisufide protects against oxygen glucose deprivation -induced apoptosis by scavenging free radicals via the PI3K/Akt -mediated Nrf2/HO-1 signaling pathway in B35 neural cells
1
2015
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Hollow Prussian Blue Nanozymes Drive Neuroprotection against Ischemic Stroke via Attenuating Oxidative Stress, Counteracting Inflammation, and Suppressing Cell Apoptosis
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Geniposide protects against hypoxia/reperfusion-induced blood-brain barrier impairment by increasing tight junction protein expression and decreasing inflammation, oxidative stress, and apoptosis in an in vitro system
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Curcumin by down-regulating NF-kB and elevating Nrf2, reduces brain edema and neurological dysfunction after cerebral I/R
1
2016
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Hexahydrocurcumin protects against cerebral ischemia/reperfusion injury, attenuates inflammation, and improves antioxidant defenses in a rat stroke model
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Sirt3 confers protection against neuronal ischemia by inducing autophagy: Involvement of the AMPK-mTOR pathway
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
ARRB1/beta-arrestin-1 mediates neuroprotection through coordination of BECN1-dependent autophagy in cerebral ischemia
1
2014
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Vitexin reverses the autophagy dysfunction to attenuate MCAO-induced cerebral ischemic stroke via mTOR/Ulk1 pathway
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Silibinin Prevents Autophagic Cell Death upon Oxidative Stress in Cortical Neurons and Cerebral Ischemia-Reperfusion Injury
1
2016
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Folic acid deficiency increases brain cell injury via autophagy enhancement after focal cerebral ischemia
1
2016
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Beneficial effect of astragalosides on stroke condition using PC12 cells under oxygen glucose deprivation and reperfusion
1
2014
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Isoquercetin attenuates oxidative stress and neuronal apoptosis after ischemia/reperfusion injury via Nrf2-mediated inhibition of the NOX4/ROS/NF-kappaB pathway
1
2018
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Isoquercetin Ameliorates Cerebral Impairment in Focal Ischemia Through Anti-Oxidative, Anti-Inflammatory, and Anti-Apoptotic Effects in Primary Culture of Rat Hippocampal Neurons and Hippocampal CA1 Region of Rats
1
2017
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Epicatechin Gallate Protects HBMVECs from Ischemia/Reperfusion Injury through Ameliorating Apoptosis and Autophagy and Promoting Neovascularization
1
2019
... Moreover, the anti-OS and anti-inflammatory effects are beneficial for post-ischemic treatment. Schizandrin A inhibits inflammation and OS through the AMPK/Nrf2 pathway [172]. Tryptanthrin [173], 3, 14, 19-triacetyl andrographolide [174], quercetin [175], diosgenin [176] and irisin [177] reduce the production of proinflammatory cytokines by inhibiting the NF-κB-related signaling pathways and suppressing ROS generation. In addition, the Zinc ion chelating agents, TPEN [178], N-acetyl lysyltyrosylcysteine amide [179], Tanshinone IIA [180], resveratrol [181] and EPO-cyclosporine combination therapy [182] preserve neuronal function through anti-OS and anti-inflammatory mechanisms of action. Furthermore, many antioxidants have effects on neuroprotection via anti-OS and anti-apoptosis. Rhein [183], deuterohemin His peptide-6 [184], acteoside [185], Radix scrophulariae [186], pomalidomide [187], and Clostridium butyricum [188] protect the brain against cerebral I/R injury by inhibiting apoptosis and OS. Similarly, adiponectin [189], YiQiFuMai [190], epigallocatechin-3-gallate (EGCG) [191], Withania somnifera [192], SMXZF [193], and diallyl trisulfide [194] lessen neuronal impairment by attenuating mitochondrial caspase-independent apoptosis. Besides, plumbagin [187], hollow prussian blue nanozymes [195], geniposide [196], curcumin [197], and hexahydrocurcumin [198] play a role in neuronal protection by anti-OS, anti-inflammatory, and anti-apoptotic activities. In addition, as autophagy is a double-edged sword, promoting or inhibiting autophagy to exert neuroprotective effects depends on different conditions. On one hand, Sirt3 [199] and β-arrestin-1 [200] promote autophagy to play a neuroprotective role. On the other hand, vitexin [201], silibinin [202]. 3-methyladenine [203], and astragalosides [204] inhibit autophagy to protect the brain. It is also worth mentioning that isoquercetin [205, 206] and EGCG [207] alleviate brain impairment via anti-OS, anti-inflammatory, anti-apoptotic, and anti-autophagocytic properties. ...
Short-term green tea supplementation prevents recognition memory deficits and ameliorates hippocampal oxidative stress induced by different stroke models in rats
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Green Tea and Red Tea from Camellia sinensis Partially Prevented the Motor Deficits and Striatal Oxidative Damage Induced by Hemorrhagic Stroke in Rats
1
2018
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Cofilin Knockdown Attenuates Hemorrhagic Brain Injury-induced Oxidative Stress and Microglial Activation in Mice
1
2018
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Exploration of MST1-Mediated Secondary Brain Injury Induced by Intracerebral Hemorrhage in Rats via Hippo Signaling Pathway
1
2019
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Dexmedetomidine Protects Against Neurological Dysfunction in a Mouse Intracerebral Hemorrhage Model by Inhibiting Mitochondrial Dysfunction-Derived Oxidative Stress
1
2019
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Oleuropein protects intracerebral hemorrhage-induced disruption of blood-brain barrier through alleviation of oxidative stress
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Overexpression of adiponectin alleviates intracerebral hemorrhage-induced brain injury in rats via suppression of oxidative stress
1
2018
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Isoliquiritigenin alleviates early brain injury after experimental intracerebral hemorrhage via suppressing ROS- and/or NF-kappaB-mediated NLRP3 inflammasome activation by promoting Nrf2 antioxidant pathway
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Sirt3 Ameliorates Oxidative Stress and Mitochondrial Dysfunction After Intracerebral Hemorrhage in Diabetic Rats
1
2018
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Sodium Benzoate Attenuates Secondary Brain Injury by Inhibiting Neuronal Apoptosis and Reducing Mitochondria-Mediated Oxidative Stress in a Rat Model of Intracerebral Hemorrhage: Possible Involvement of DJ-1/Akt/IKK/NFkappaB Pathway
1
2019
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Carnosine Attenuates Brain Oxidative Stress and Apoptosis After Intracerebral Hemorrhage in Rats
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Metformin Attenuates Neurological Deficit after Intracerebral Hemorrhage by Inhibiting Apoptosis, Oxidative Stress and Neuroinflammation in Rats
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Baicalein Promotes Neuronal and Behavioral Recovery After Intracerebral Hemorrhage Via Suppressing Apoptosis, Oxidative Stress and Neuroinflammation
1
2017
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Neuroprotective effects of hydrogen inhalation in an experimental rat intracerebral hemorrhage model
1
2018
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Protocatechuic acid exerts protective effects via suppression of the P38/JNK- NF-kappaB signalling pathway in an experimental mouse model of intracerebral haemorrhage
1
2019
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Therapeutic targeting of oxygen-sensing prolyl hydroxylases abrogates ATF4-dependent neuronal death and improves outcomes after brain hemorrhage in several rodent models
1
2016
... Like ischemic stroke, antioxidant defenses involving OS and OS-related processes are also found in ICH and SAH. Green tea [208, 209], cofilin [210], and mammalian sterile 20-like kinase-1 [211] play a major role in ICH-induced SBI. Melatonin [51] and dexmedetomidine [212] reduce mitochondrial impairments and ameliorate SBI. Analogously, oleuropein [213] and adiponectin [214] attenuate brain edema and preserve the BBB structure in a dose-dependent manner. In addition, isoliquiritigenin [215] and Sirt3 [216] improve ICH by inhibiting inflammatory and OS activities, whereas Sodium Benzoate [217] and carnosine [218] attenuate SBI and brain edema through suppression of neuronal apoptosis. As such, metformin [219], baicalein [220], hydrogen gas [221], protocatechuic acid [222], and hypoxia-inducible factor prolyl hydroxylase domain metalloenzymes [223] prevent neurological deficits after ICH by inhibiting apoptosis, OS, and neuroinflammation in rats. ...
Dimethylfumarate alleviates early brain injury and secondary cognitive deficits after experimental subarachnoid hemorrhage via activation of Keap1-Nrf2-ARE system
1
2015
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Telmisartan ameliorates oxidative stress and subarachnoid haemorrhage-induced cerebral vasospasm
1
2016
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Nebivolol attenuates cerebral vasospasm both by increasing endothelial nitric oxide and by decreasing oxidative stress in an experimental subarachnoid haemorrhage
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Curcumin mitigates cerebral vasospasm and early brain injury following subarachnoid hemorrhage via inhibiting cerebral inflammation
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Pterostilbene Attenuates Early Brain Injury Following Subarachnoid Hemorrhage via Inhibition of the NLRP3 Inflammasome and Nox2-Related Oxidative Stress
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Apigenin attenuates oxidative stress and neuronal apoptosis in early brain injury following subarachnoid hemorrhage
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Peroxiredoxin 1/2 protects brain against H2O2-induced apoptosis after subarachnoid hemorrhage
1
2019
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Docosahexaenoic Acid Alleviates Oxidative Stress-Based Apoptosis Via Improving Mitochondrial Dynamics in Early Brain Injury After Subarachnoid Hemorrhage
1
2018
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Hydrogen Sulfide Ameliorates Early Brain Injury Following Subarachnoid Hemorrhage in Rats
1
2016
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Cysteamine alleviates early brain injury via reducing oxidative stress and apoptosis in a rat experimental subarachnoid hemorrhage model
1
2015
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Effect of Gastrodin on Early Brain Injury and Neurological Outcome After Subarachnoid Hemorrhage in Rats
1
2019
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Progesterone alleviates acute brain injury via reducing apoptosis and oxidative stress in a rat experimental subarachnoid hemorrhage model
1
2015
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
AVE 0991 attenuates oxidative stress and neuronal apoptosis via Mas/PKA/CREB/UCP-2 pathway after subarachnoid hemorrhage in rats
1
2019
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Memantine alleviates brain injury and neurobehavioral deficits after experimental subarachnoid hemorrhage
1
2015
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Cerebroprotection by salvianolic acid B after experimental subarachnoid hemorrhage occurs via Nrf2- and SIRT1-dependent pathways
1
2018
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Sirtuin 1 activation protects against early brain injury after experimental subarachnoid hemorrhage in rats
1
2016
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Salvianolic acid A attenuates early brain injury after subarachnoid hemorrhage in rats by regulating ERK/P38/Nrf2 signaling
1
2017
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Allicin attenuates early brain injury after experimental subarachnoid hemorrhage in rats
1
2019
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Mitophagy Reduces Oxidative Stress Via Keap1 (Kelch-Like Epichlorohydrin-Associated Protein 1)/Nrf2 (Nuclear Factor-E2-Related Factor 2)/PHB2 (Prohibitin 2) Pathway After Subarachnoid Hemorrhage in Rats
1
2019
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Melatonin-enhanced autophagy protects against neural apoptosis via a mitochondrial pathway in early brain injury following a subarachnoid hemorrhage
1
2014
... Many preclinical experiments in SAH have demonstrated that inhibiting OS and OS-mediated pathological processes alleviates alleviates EBI, SBI, and neuronal deficits. Dimethyl formamide improves cognitive dysfunction via the Keap1-Nrf2-ARE system [224]. Telmisartan [225], nebivolol [226] and curcumin [227] can ameliorate cerebral vasospasm, and Pterostilbene can attenuate EBI by inhibiting the NLRP3 inflammasome [228]. Some antioxidants (e.g., apigenin [229], peroxiredoxin1/2 [230], docosahexaenoic acid [231], sodium hydrosulfide [232], cysteamine [233], gastrodin [234], naringin [60], and progesterone [235]) play a neuroprotective role in EBI through the effects of anti-apoptosis after SAH, but other antioxidants (e.g. AVE 0991 and mangiferin [56, 236] memantine [59, 237], salvianolic acid B [238, 239], salvianolic acid A [240], and allicin [241]) protect against SAH-induced oxidative injury via inhibition of oxidative, inflammatory, and apoptotic pathways. Stimulating autophagy to inhibit apoptosis of neural cells also works in improving neurological outcome with administration of mitoquinone [242] and melatonin [243]. ...
Inhibition of phosphodiesterase 2 reverses gp91phox oxidase-mediated depression- and anxiety-like behavior
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
4,4'-Dichlorodiphenyl diselenide reverses a depressive-like phenotype, modulates prefrontal cortical oxidative stress and dysregulated glutamatergic neurotransmission induced by subchronic dexamethasone exposure to mice
1
2019
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Homocysteine aggravates ROS-induced depression of transmitter release from motor nerve terminals: potential mechanism of peripheral impairment in motor neuron diseases associated with hyperhomocysteinemia
1
2015
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Vitamin D Supplementation Affects the Beck Depression Inventory, Insulin Resistance, and Biomarkers of Oxidative Stress in Patients with Major Depressive Disorder: A Randomized, Controlled Clinical Trial
1
2016
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
2,3,5,4'-Tetrahydroxystilbene-2-O-beta-D-glucoside Reverses Stress-Induced Depression via Inflammatory and Oxidative Stress Pathways
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Antidepressant activity of vorinostat is associated with amelioration of oxidative stress and inflammation in a corticosterone-induced chronic stress model in mice
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Naringenin protects against oxido-inflammatory aberrations and altered tryptophan metabolism in olfactory bulbectomized-mice model of depression
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Naringenin attenuates behavioral derangements induced by social defeat stress in mice via inhibition of acetylcholinesterase activity, oxidative stress and release of pro-inflammatory cytokines
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Activation of ATP-sensitive potassium channel by iptakalim normalizes stress-induced HPA axis disorder and depressive behaviour by alleviating inflammation and oxidative stress in mouse hypothalamus
1
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Silymarin ameliorates experimentally induced depressive like behavior in rats: Involvement of hippocampal BDNF signaling, inflammatory cytokines and oxidative stress response
1
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Protective effects of resveratrol on mitochondrial function in the hippocampus improves inflammation-induced depressive-like behavior
1
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Honokiol abrogates lipopolysaccharide-induced depressive like behavior by impeding neuroinflammation and oxido-nitrosative stress in mice
1
2014
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Ketamine's antidepressant effect is mediated by energy metabolism and antioxidant defense system
2
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
... ] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Postnatal melatonin treatment protects against affective disorders induced by early-life immune stimulation by reducing the microglia cell activation and oxidative stress
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Oxytocin improves animal behaviors and ameliorates oxidative stress and inflammation in autistic mice
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Effects of vanillin on potassium bromate-induced neurotoxicity in adult mice: impact on behavior, oxidative stress, genes expression, inflammation and fatty acid composition
1
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Trigonelline mitigates lipopolysaccharide-induced learning and memory impairment in the rat due to its anti-oxidative and anti-inflammatory effect
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Quercetin prevents chronic unpredictable stress induced behavioral dysfunction in mice by alleviating hippocampal oxidative and inflammatory stress
1
2017
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
alpha-Tocopherol Ameliorates Redox Equilibrium and Reduces Inflammatory Response Caused by Chronic Variable Stress
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Baicalin ameliorates chronic mild stress-induced depression-like behaviors in mice and attenuates inflammatory cytokines and oxidative stress
1
2019
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
The selenium-containing compound 3-((4-chlorophenyl)selanyl)-1-methyl-1H-indole reverses depressive-like behavior induced by acute restraint stress in mice: modulation of oxido-nitrosative stress and inflammatory pathway
1
2019
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
The effects of ketamine on prefrontal glutamate neurotransmission in healthy and depressed subjects
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Uncoupling protein 2 modulation of the NLRP3 inflammasome in astrocytes and its implications in depression
1
2016
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Dl-3-n-Butylphthalide improves lipopolysaccharide-induced depressive-like behavior in rats: involvement of Nrf2 and NF-kappaB pathways
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Effects of indole-3-carbinol on clonidine-induced neurotoxicity in rats: Impact on oxidative stress, inflammation, apoptosis and monoamine levels
1
2014
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Effect of 25-methoxy hispidol A isolated from Poncirus trifoliate against bacteria-induced anxiety and depression by targeting neuroinflammation, oxidative stress and apoptosis in mice
1
2019
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Allicin attenuated chronic social defeat stress induced depressive-like behaviors through suppression of NLRP3 inflammasome
1
2019
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Apocynum venetum leaf extract reverses depressive-like behaviors in chronically stressed rats by inhibiting oxidative stress and apoptosis
1
2018
... Apart from stroke, using antioxidants may help improve depression. Generally, antioxidants exert antidepressant effects through their anti-OS characteristics (Table 2). Bay 60-7550 [244], p-chloro-diphenyl diselenide [245], homocysteine [246], and vitamin D [247] can attenuate depressive-like behaviors and improve depression. In a sense, the latest studies have focused on antioxidants that act against OS-induced inflammation. Examples of this include 2,3,5,4'-tetrahydroxystilbene-2-O-β-D- gluco-pyranoside [248] and vorinostat [249], which ameliorate inflammatory damage and OS to exert the antidepressant effect. Moreover, naringenin [250, 251], iptakalim [252], silymarin [253], resveratrol [254], honokiol [255], ketamine [256], melatonin [257], oxytocin [258], vanillin [259], trigonelline [260], quercetin [261], α-tocopherol [262], baicalin [263] and selenium-containing compounds [264] have neuroprotective effects in the hippocampus of mice due to their antioxidant and anti-inflammatory properties. Particularly, ketamine is involved in inducing the rapid antidepressant effects through increasing the release of glutamate in the body [265] and affecting energy metabolism in MDD [256]. Furthermore, some antioxidants with multifunctional properties demonstrate anti-OS, anti-inflammatory, and anti-apoptotic characteristics in depression. These antioxidants include mitochondrial uncoupling protein 2 [266], dl-3-n-butylphthalide [267], indole-3-carbinol [268], or 25-methoxyhispidol A [269] and allicin [270]. Of note, AVLE treatment has similar effects compared to fluoxetine on depression in the rat hippocampus [271]. ...
Dietary flavonoid intake and the risk of stroke: a dose-response meta-analysis of prospective cohort studies
1
2016
... In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276]. ...
Protective Effect of Quercetin against Oxidative Stress-Induced Cytotoxicity in Rat Pheochromocytoma (PC-12) Cells
1
2017
... In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276]. ...
Gaultheria trichophylla (Royle): a source of minerals and biologically active molecules, its antioxidant and anti-lipoxygenase activities
1
2017
... In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276]. ...
Inhibition of Human Monoamine Oxidase: Biological and Molecular Modeling Studies on Selected Natural Flavonoids
1
2016
... In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276]. ...
Dietary flavonoid intake and risk of incident depression in midlife and older women
1
2016
... In stroke, high dietary flavonoid intake may modestly lower the risk of stroke due to its role in constraining OS-induced mitochondrial lipid peroxidation [272]. In depression, flavonoids can limit ROS production and promote the chelation of transition metal elements [273, 274]. Also, flavonoids act as reversible and competitive human monoamine oxidase inhibitors in the CNS, leading to increased central neurotransmission [275]. Moreover, a study enrolling 10,752 depressed individuals shows that higher flavonoid intake may be linked to lower depression risk, especially among older women [276]. ...
Safety and efficacy of uric acid in patients with acute stroke (URICO-ICTUS): a randomised, double-blind phase 2b/3 trial
1
2014
... UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281]. ...
Uric Acid Therapy Improves Clinical Outcome in Women With Acute Ischemic Stroke
1
2015
... UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281]. ...
Uric acid improves glucose-driven oxidative stress in human ischemic stroke
1
2015
... UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281]. ...
Association between the antioxidant uric acid and depression and antidepressant medication use in 96 989 individuals
1
2017
... UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281]. ...
Antioxidant uric acid in treated and untreated subjects with major depressive disorder: a meta-analysis and meta-regression
1
2018
... UA is the primary endogenous antioxidant in blood. A randomized, double-blind phase 2b/3 trial (URICOI-CTUS) indicates that UA is safe without any concerns regarding safety, although the addition of UA to thrombolytic therapy does not raise the proportion of patients with excellent outcome after stroke [277]. However, a reanalysis of the URICOICTUS trial suggests that UA suppresses infarct growth, and is more effective than the placebo in reaching an excellent outcome in patients who are treated with alteplase following acute ischemic stroke [278]. Another clinical study supports the efficacy of UA therapy by showing decreased infarct growth and enhanced outcome in stroke survivors [279]. In depression, researchers have found that UA is associated with a low risk of hospitalization for depression after examining the plasma levels of UA in 96,989 depressed subjects [280]. Moreover, the efficacy of UA in depressed patients is supported by another large-scale study [281]. ...
Melatonin use for neuroprotection in perinatal asphyxia: a randomized controlled pilot study
1
2015
... To our knowledge, few clinical trials have studied the neuroprotective effects of melatonin on cerebral ischemia prevention. A prospective trial involving 45 neonates indicates that it is practicable to provide early administration of melatonin to ameliorate brain injury in a choking newborn infant [282]. In regards to treatment of MDD, preliminary findings of a study reveal that a combination of buspirone with melatonin can partially benefit cognitive function [283]. ...
Changes in cognitive symptoms after a buspirone-melatonin combination treatment for Major Depressive Disorder
1
2015
... To our knowledge, few clinical trials have studied the neuroprotective effects of melatonin on cerebral ischemia prevention. A prospective trial involving 45 neonates indicates that it is practicable to provide early administration of melatonin to ameliorate brain injury in a choking newborn infant [282]. In regards to treatment of MDD, preliminary findings of a study reveal that a combination of buspirone with melatonin can partially benefit cognitive function [283]. ...
Post-stroke depression: Mechanisms and pharmacological treatment
1
2018
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Effects of fluoxetine on functional outcomes after acute stroke (FOCUS): a pragmatic, double-blind, randomised, controlled trial
1
2019
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Comparative efficacy and acceptability of antidepressant treatment in poststroke depression: a multiple-treatments meta-analysis
1
2017
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Fluoxetine for motor recovery after acute ischaemic stroke (FLAME): a randomised placebo-controlled trial
1
2011
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Management of post-stroke mood and emotional disturbances
1
2017
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Efficacy of early administration of escitalopram on depressive and emotional symptoms and neurological dysfunction after stroke: a multicentre, double-blind, randomised, placebo-controlled study
1
2017
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Escitalopram and problem-solving therapy for prevention of poststroke depression: a randomized controlled trial
1
2008
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Efficacy of Citalopram on Acute Ischemic Stroke Outcome: A Randomized Clinical Trial
1
2017
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
TALOS: a multicenter, randomized, double-blind, placebo-controlled trial to test the effects of citalopram in patients with acute stroke
1
2015
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Impact of prestroke selective serotonin reuptake inhibitor treatment on stroke severity and mortality
1
2014
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Prevention of poststroke depression with milnacipran in patients with acute ischemic stroke: a double-blind randomized placebo-controlled trial
1
2011
... At present, antidepressant drugs are widely used for treating post-stroke depression (PSD), but there are many adverse reactions at the same time. These events include gastroenterological symptoms, falls/fracture, and epilepsy [284]. Therefore, future clinical trials should emphasize their focus on the efficacy and safety of antidepressant drugs in preventing or treating stroke (Table 4). Herein, we summarize some clinical trials in recent years with the objective of determining whether antidepressants (e.g., fluoxetine, escitalopram, milnacipran) are necessary for PSD patients. Results of FOCUS (effects of fluoxetine on functional outcomes after acute stroke) represent a lower incidence of new depression and a higher risk of bone fractures in the group in which fluoxetine is allocated, when compared with the controls. Those outcomes do not support the routine use of fluoxetine in preventing PSD or promoting function recovery [285]. In addition, a meta-analysis of 12 suitable trials demonstrated that fluoxetine is the worst choice for PSD treatment, while paroxetine is an effective and acceptable drug [286]. However, FLAME (fluoxetine for motor recovery after acute ischemic stroke) exhibits a positive connection between motor recovery and fluoxetine use in PSD patients [287]. The second drug is escitalopram. Kim K et al. did a Cochrane systematic review of 13 trials of antidepressant drugs, and confirmed that escitalopram is the most tolerated SSRI [288]. Another study also indicates that escitalopram was well-tolerated, though it does not have any effects on the depressive symptoms in PSD patients [289]. Robinson et al. illustrated that escitalopram is significantly effective at decreasing the incidence of depression in patients with recent stroke over 12-month therapy [290]. Thirdly, citalopram is reported to be a safe medication in patients with acute ischemic stroke [291], and it improves PSD [292]. In conclusion, fluoxetine is not necessary for PSD treatment, but it is good for motor recovery. Escitalopram and citalopram are both tolerated and may have benefits in PSD patients. However, the question of whether to allocate antidepressants in post-stroke patients is still controversial, when considering efficacy and safety. Additionally, is pre-stroke SSRI therapy useful for patients with stroke? Mortensen et al. conducted a study enrolling hemorrhagic stroke patients and ischemic stroke patients. They found that pre-stroke SSRI use increased severity and mortality in patients with hemorrhagic stroke, but there was no increase in ischemic stroke patients [293]. Finally, milnacipran also plays a role in preventing PSD and it is safe to use, without any serious adverse events, according to an RCT performed by Ching-Shu Tsai [294]. Due to the limited data showing a relationship between depressants and PSD, more multicenter clinical trials should be conducted. ...
Neuropsychiatric sequelae of stroke
1
2016
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Depression predictors within six months of ischemic stroke: The DEPRESS Study
1
2016
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Blood-Brain Barrier: From Physiology to Disease and Back
1
2019
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Dietary flavonoid intake and the risk of stroke: a dose-response meta-analysis of prospective cohort studies
1
2016
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Melatonin as a treatment for mood disorders: a systematic review
1
2017
... This review discusses the central role of OS in the “aging-stroke-depression” network and antioxidants in treating stroke or depression, as well as antidepressant in PSD. Although all of the studies have immeasurable contributions to scientific exploration, there are some limitations in the current studies. First, depression is a psychiatric disorder lacking effective biomarkers or methods, and depressive symptoms may be less apparent, making it difficult to procure an accurate diagnosis for depression [10]. Relatively uniform and accurate diagnostic criteria for depression must be improved [295]. Second, this review focuses on major depression, leaving an unclear relationship between other depression phases and stroke. Third, although this review suggests aging as a risk for stroke and depression, the link is often found in aged adults, but not analyzed in other age groups [296, ]33]. For future studies, it is suggested that there should be an emphasis on certain age demographics. Fourth, human brains have a natural and complex protective barrier, which prevents antioxidant drugs from entering into the CNS, partially explaining the reduced efficacy of these drugs in experimental models [297]. Finding carriers that can penetrate the blood-brain barrier, such as nanoparticles, may address this problem. Fifth, there are data deviations due to the lack of dose-response analysis, limitations on sample size, representativeness of the sample, statistical methods, inclusion criteria, and follow-up [37, 298, 299]. Therefore, studies should be conducted in a more precise manner. Furthermore, drug dose, window time, and methods of administration also warrant our attention. All things considered, further studies are required to improve the diagnosis of depression, to find drugs directed at overproduced ROS with higher efficacy and safety, and to enhance the quality of life after diseases. ...
Free radicals and antioxidants in normal physiological functions and human disease
1
2007
... Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303]. ...
Reactive Oxygen Species in Metabolic and Inflammatory Signaling
1
2018
... Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303]. ...
NOX4 activity is determined by mRNA levels and reveals a unique pattern of ROS generation
1
2007
... Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303]. ...
Pharmacological applications of antioxidants: lights and shadows
1
2014
... Schematic model of the main source of ROS and redox reaction. ROS are generated mainly from enzymatic reactions in the cytoplasm, endoplasmic reticulum, mitochondria, and peroxisome [300]. Specifically, overproduced mitoROS can affect metabolic pathways, such as alteration of protein translation, oxidation of lipid and DNA, and impairment of ATP synthesis [301]. Moreover, assembled NOX (NOX1 and NOX2) complex transports an electron from cytosolic NADPH to oxygen to form superoxide on the extracellular side [13]. The NOX4 complex rapidly converts the superoxide to H2O2, which undergoes a Fenton reaction to produce hydroxyl radicals and ions, and to regulate many downstream effects [302]. However, these oxidative events are inhibited by antioxidants, such as SOD and CAT/GPx. Activation of Nrf2-ARE pathway increases antioxidants, such as HO-1, SOD1, and CAT to protect cells from FR accumulation [303]. ...
Inflammatory Pathways Following Subarachnoid Hemorrhage
1
2019
... Pathogenesis and correlation between stroke and depression. There are different mechanisms in ischemic stroke [16], ICH [17], SAH [304], and depression [10]. Stroke and depression are associated with oxidative stress. Due to overactive OS activity and impaired anti-OS defenses, 16-30% of ischemic stroke survivors [31, 32], 25% of ICH [33] patients, and 50% of SAH [34] patients may develop depression later, but the age groups vary among studies. Conversely, depression increases the risk of stroke by 33% in patients who experience stressful life events [37]. ...
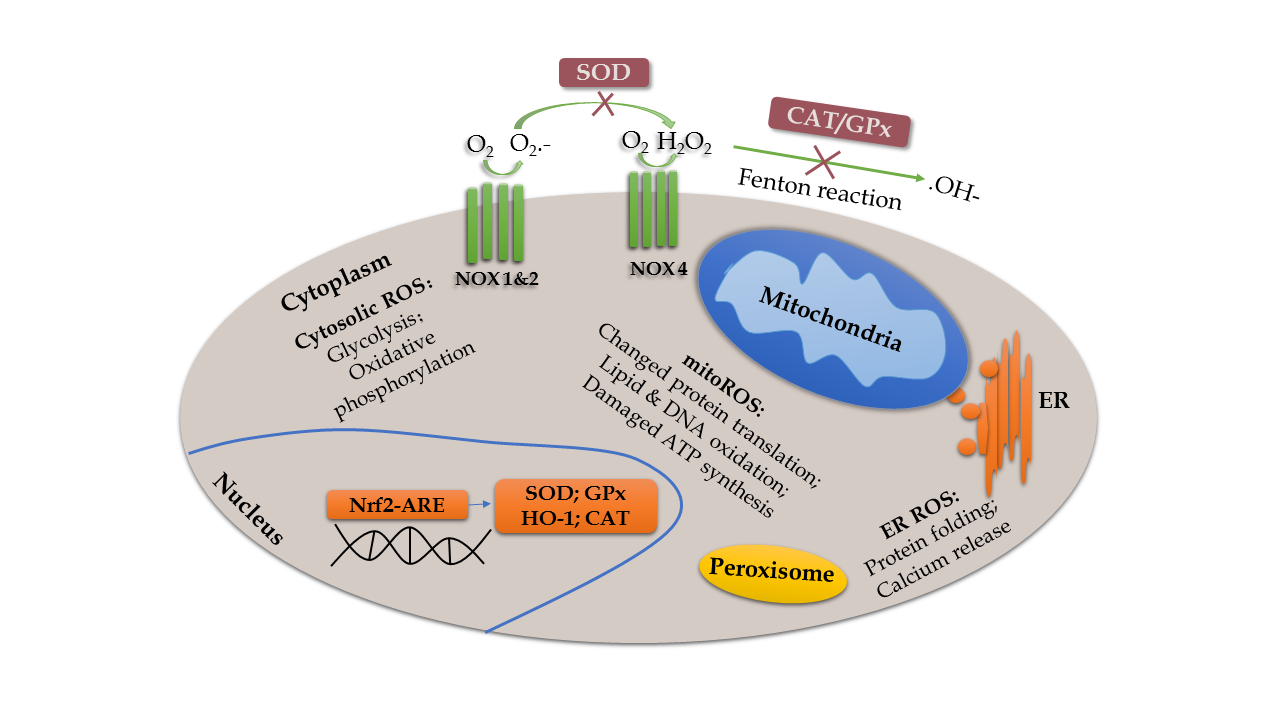
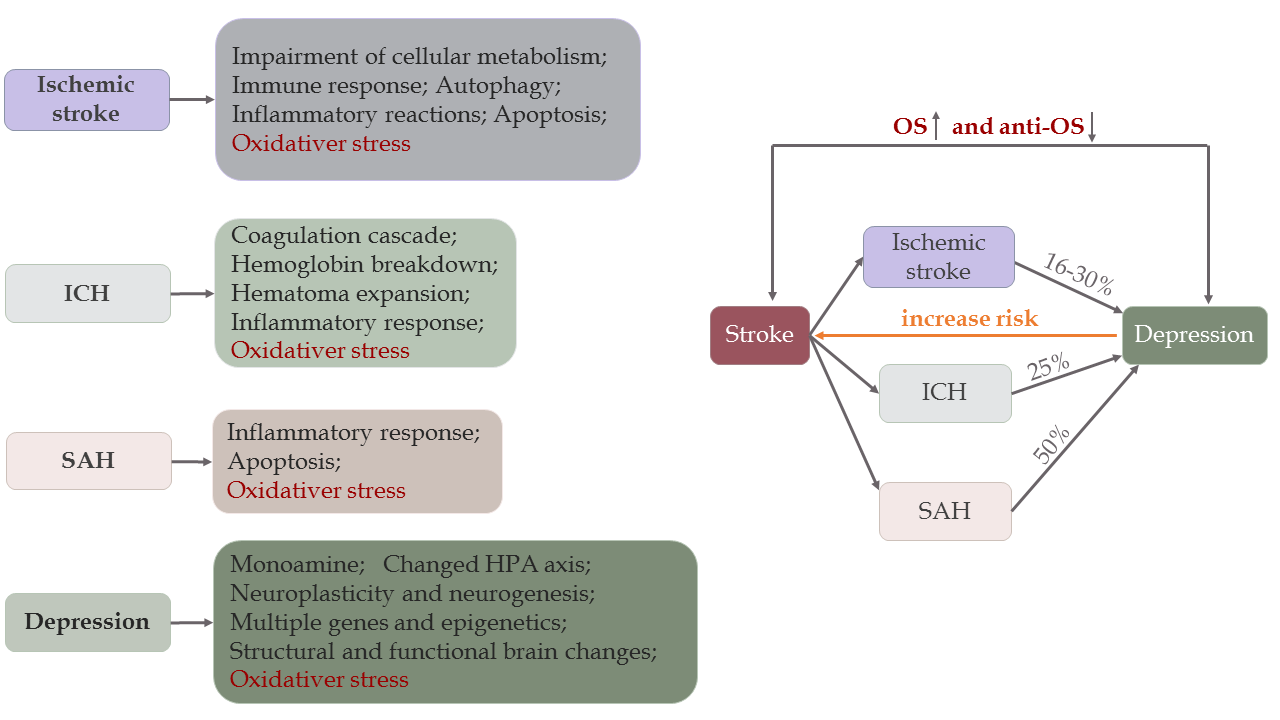
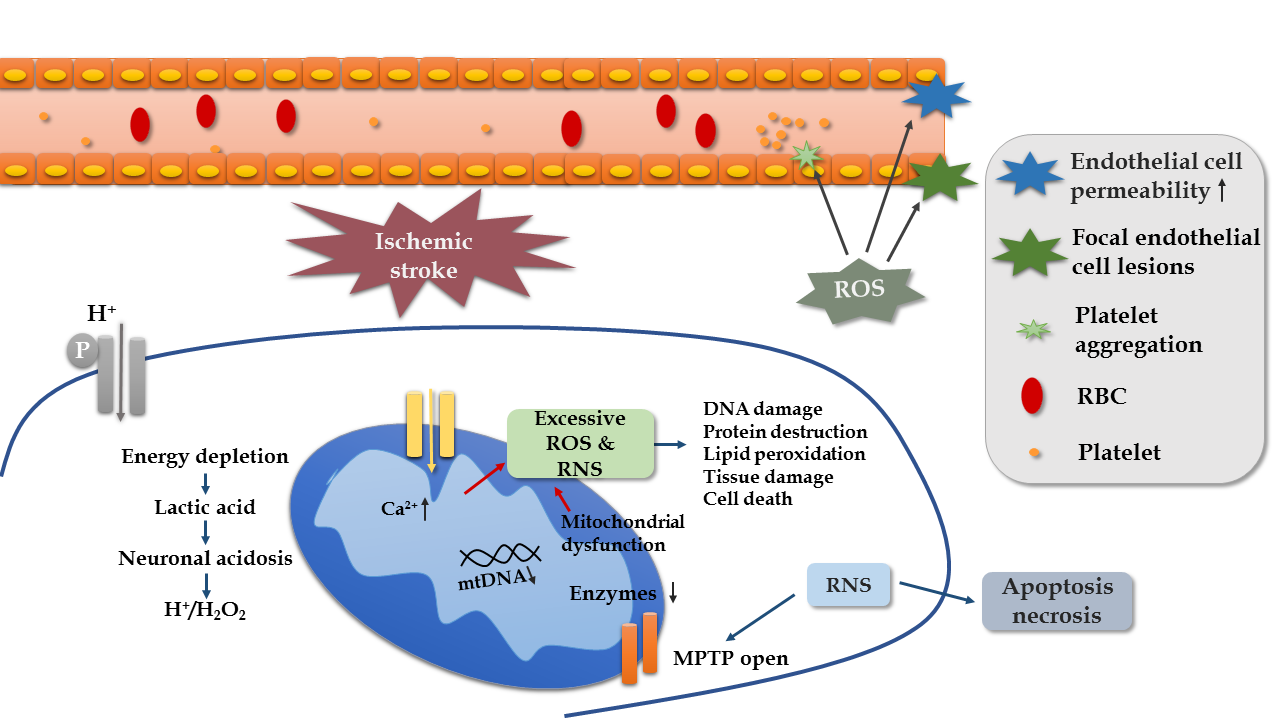
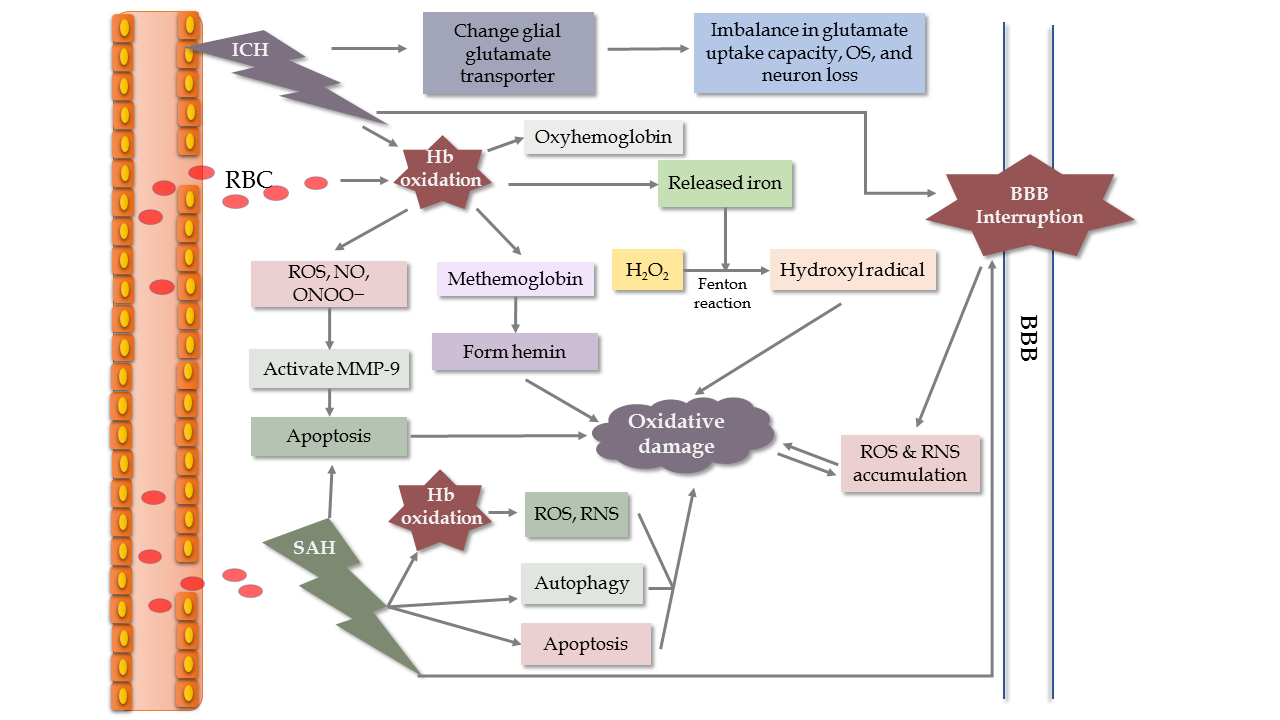
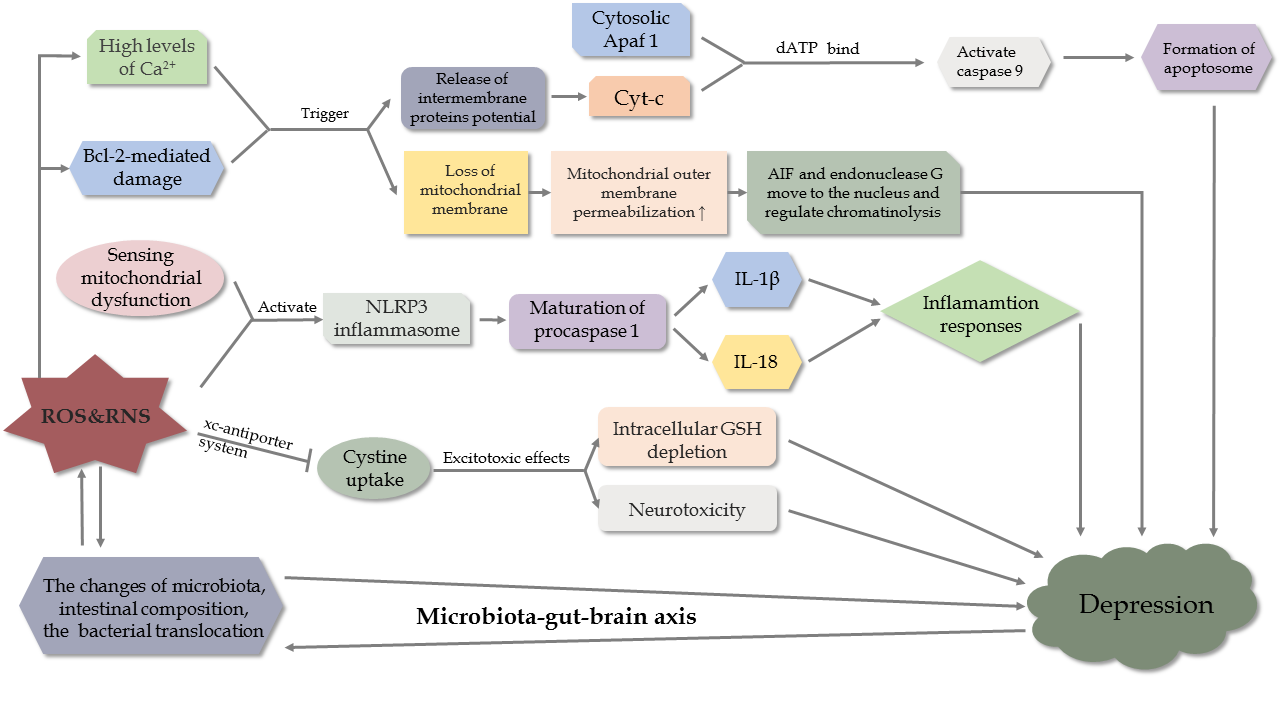

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



