COVID-19 Virulence in Aged Patients Might Be Impacted by the Host Cellular MicroRNAs Abundance/Profile
Fulzele Sadanand1, 2, *, Sahay Bikash3, Yusufu Ibrahim1, Lee Tae Jin4, Sharma Ashok4, Kolhe Ravindra5, Isales Carlos M1, 2
1Department of Medicine, Augusta University, Augusta, GA, USA.2Center for Healthy Aging, Augusta University, Augusta, GA, USA.3Department of Infectious Diseases and Immunology, University of Florida, Gainesville, FL, USA.4Center for Biotechnology and Genomic Medicine, Augusta University, Augusta, GA 30912, USA.5Departments of Pathology, Augusta University, Augusta, GA 30912, USA
通讯作者: Correspondence should be addressed to: Dr. Sadanand Fulzele, Department of Medicine, Center for Healthy Aging, Augusta University, Augusta, GA 30912, USA. Email: SFULZELE@augusta.edu.Correspondence should be addressed to: Dr. Sadanand Fulzele, Department of Medicine, Center for Healthy Aging, Augusta University, Augusta, GA 30912, USA. Email: SFULZELE@augusta.edu.
收稿日期:2020-04-1
修回日期: 2020-04-27
接受日期: 2020-04-28
网络出版日期: 2020-05-09
版权声明:
2020 This is an Open Access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
展开
Abstract
The World health organization (WHO) declared Coronavirus disease 2019 (COVID-19) a global pandemic and a severe public health crisis. Drastic measures to combat COVID-19 are warranted due to its contagiousness and higher mortality rates, specifically in the aged patient population. At the current stage, due to the lack of effective treatment strategies for COVID-19 innovative approaches need to be considered. It is well known that host cellular miRNAs can directly target both viral 3'UTR and coding region of the viral genome to induce the antiviral effect. In this study, we did in silico analysis of human miRNAs targeting SARS (4 isolates) and COVID-19 (29 recent isolates from different regions) genome and correlated our findings with aging and underlying conditions. We found 848 common miRNAs targeting the SARS genome and 873 common microRNAs targeting the COVID-19 genome. Out of a total of 848 miRNAs from SARS, only 558 commonly present in all COVID-19 isolates. Interestingly, 315 miRNAs are unique for COVID-19 isolates and 290 miRNAs unique to SARS. We also noted that out of 29 COVID-19 isolates, 19 isolates have identical miRNA targets. The COVID-19 isolates, Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) showed six, four, and four unique miRNAs targets, respectively. Furthermore, GO, and KEGG pathway analysis showed that COVID-19 targeting human miRNAs involved in various age-related signaling and diseases. Recent studies also suggested that some of the human miRNAs targeting COVID-19 decreased with aging and underlying conditions. GO and KEGG identified impaired signaling pathway may be due to low abundance miRNA which might be one of the contributing factors for the increasing severity and mortality in aged individuals and with other underlying conditions. Further, in vitro and in vivo studies are needed to validate some of these targets and identify potential therapeutic targets.
FulzeleSadanand, SahayBikash, YusufuIbrahim, LeeTae Jin, SharmaAshok, KolheRavindra, IsalesCarlos M. COVID-19 Virulence in Aged Patients Might Be Impacted by the Host Cellular MicroRNAs Abundance/Profile[J]. Aging and Disease, 2020, 11(3): 509-522 https://doi.org/10.14336/AD.2020.0428
Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7].
The World Health Organization (WHO) reported 2,920,905 confirmed infected, and 203,269 COVID-19 related deaths globally as of April 25th, 2020 [8]. Drastic measures to combat SARS-CoV-2 are warranted due to its contagiousness, viability, and higher death toll compared to its predecessor SARS-CoV. Although SARS-CoV had a significantly higher case-fatality (around 10%) [9], the virus was not viable enough to remain in the human population and never spread in the United States and other countries like SARS-CoV-2 [10]. SARS-CoV-2, on the other hand, appears to rapidly spread, which is contributing to its global spread and a significantly higher number of cases.
Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16].
MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets.
Table 1
Table 1 Details of SARS and COVID-19 isolates from different geographic locations, sequence length, and the number of human miRNA targets.
Viral genome sequence retrieval, homology, and phylogenetic analyses
The complete genome sequences of the SARS and COVID-19 isolates were retrieved from the GenBank database. We retrieved four SARS and 29 COVID-19 sequences from NCBI and GISAID for for homology and phylogenetic analysis. Details of sequence identification are summarizing in Table 1. The sequences were aligned using the Multiple Sequence Alignment tool at Clustal Omega 1.2.3 on Geneious Prime 2020.1.1. The phylogenetic alignment tree generated using the neighbor-joining method.
COVID-19 genome and human MiRNA target analysis
As previously mentioned above, miRNAs are known to target 3'UTR and coding sequences and prevent mRNA translation or degrade RNA. Keeping this in consideration, we used the whole viral genome sequence for miRNA target analysis. We used miRDB (http://www.mirdb.org/) software to identify novel human miRNAs targeting the COVID-19 viral genome [27, 28]. Furthermore, we used this data to correlate with the existing literature.
GO, and KEGG pathway analysis
Gene Ontology (GO) and KEGG signaling pathway analyses were performed on human microRNAs targeting the COVID-19 genome using DIANA-miRPath v 3.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php) [29]. We used miRNAs with a target score above 90 because these miRNAs have high probability of being real targets [27, 28].
RESULTS
Viral genome sequence homology and phylogenetic analyses
The sequences of the COVID-19 isolates have been stored, compiled, and analyzed by various sources; one of the major resources for these sequences has been the Global Initiative on Sharing All Influenza Data (GISAID), which is a public-private initiative initially made for sharing sequences of influenza data. The organization provides a complete daily analysis of the viruses uploaded by the various researchers across the Globe. The COVID-19 belongs to a family of RNA viruses, which are very stable, unlikely other RNA viruses common to us, such as Human Immunodeficiency Virus (HIV), and Foot and Mouth Diseases Virus (FMDV).
Table 2
Table 2 Sequence homology between the SARS and COVID-19 isolates from different geographic locations.
However, GISAID showed a constant drift in the COVID-19 population-based upon three specific mutations in its genome. To assimilate different sequences, we chose 29 genome sequences of COVID-19 from five continents covering 17 countries (Table 1). To gather the most diverse sequences, we collected these sequences based upon their date of isolation. Among 29 viral sequences, six isolated in 2019, and the remaining 23 were isolated at different months of 2020 from January to April (Table 1). These 29 COVID-19 sequences were compared with the four SARS genome sequences to evaluate the differences between the COVID-19 and SARS and among different isolates of COVID-19. The sequences were analyzed using Geneious Prime 2020.1.1. The data suggested the SARS genomes are approximately 78.7% similar to the COVID-19 sequences (Table 2). All 29 sequences of COVID-19 are very similar (99.9% - 100% sequence similarity). Phylogenetic analysis showed that COVID-19 is closely related to SARS isolates but still genetically different (Fig. 1). All 29 COVID-19 isolates are close to each other with little change in sequence. Since variability among different COVID-19 isolates was minimal, we calculated the number of different individual nucleotides in each COVID-19 isolates that revealed a total nucleotide difference among these COVID-19 is from 1-55 nucleotides in the entire genome of approximately 29kb (Supplementary Table 1). The most diverse sequence that has a difference of 55 nucleotides were between the isolates from Australia (MT007544.1) and Turkey (EPI_ISL_427391) isolated in 2019 and April 2020, respectively (Fig. 1 & Table 2, Supplementary Table 1).
Figure 1. Phylogenetic analysis of Coronavirus isolates from different geographic locations. The phylogenetic analysis shows sequence relatedness among COVID-19 isolates (blue) and SARS isolates (black).
Table 3
Table 3 List of human miRNAs with higher target score (above 94), the number of binding sites, and miRNAs seed binding site on COVID-19 isolates.
miRNAs
Target Score
Number of Sites and Seed locations of miRNAs and COVID-19 genome binding sites
miR-15b-5p miR-15a-5p
99
16 SITES (3163, 5384, 8458, 8614, 13090, 14562, 14781, 19857, 24094, 24634, 25683, 26723, 28921, 28935, 28938, 29023) (Note: miR-15b-5p, and miR-15a-5p have same target site)
As we mentioned above, we performed miRNA analysis on the whole genome of SARS and COVID-19 due to the efficiency of miRNAs targeting both 3'UTR and coding region. Our analysis found some interesting results. We found 848 common miRNAs targeting the SARS genome (four isolates, NC_004718.3, AY291451, AY338175, AY348314) and 873 common microRNAs targeting the 29 isolates of COVID-19 genome (Supplementary Table 2). Out of a total of 848 miRNAs from SARS, only 558 commonly present in all COVID-19 isolates (Fig. 2, Supplementary Table 2). Interestingly, 315 miRNAs are unique for COVID-19 isolates and 290 miRNAs unique to SRAS (Fig. 2). Furthermore, the COVID-19 targeting miRNAs with a higher target score (above 94) showed more than ten target sites and maximum complementary miRNA-RNA seed paring (~6-8 nucleotide seed base pairing) (Table 3). We also noted that out of 29 COVID-19 isolates, 19 have identical miRNA targets (Table 4). Ten isolates have unique miRNAs targets. Among ten isolates, Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) showed six, four, and four unique miRNAs targets respectively. The detail is given in the table (Table 4).
Table 4
Table 4 Summary of important findings on human miRNAs targeting SARS and COVID-19 genome.
Serial. No
Important findings on human miRNAs targeting Coronavirus
Figure 2. Common and different human miRNAs targeting SARS and COVID-19 isolates from different geographic locations.
GO and KEGG pathway analysis of miRNAs targeting COVID-19
To identify the functional relevance of human miRNAs targeting COVID-19, we performed Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway annotation and GO analysis. The KEGG annotation analysis data reveal that these miRNAs plays important in various Cancer signaling (e.g Renal, Pancreatic, Prostate, Colorectal, Melanoma ), Hippo signaling pathway, cardiomyopathy, Wnt signaling, Circadian rhythm, stem cell signaling, Adrenergic signaling in cardiomyocytes and number of other signaling pathways (Table 5). GO analysis data showed more than 72 biological processes were associated with the miRNAs targeting COVID-19 (Table 6). The biological processes such as organelle, ion binding, cellular, biosynthesis process, protein complex, immune response, and viral process are regulated by human miRNAs targeting the COVID-19 genome. Details of KEGG annotation and GO analysis are shown in Table 5, and Table 6, respectively.
DISCUSSION
Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31].
Table 6
Table 6 Human miRNAs targeting the COVID-19 genome regulating GO pathway.
GO Category
p-value
#genes
#miRNAs
organelle
1.26E-49
980
64
ion binding
5.53E-28
611
64
cellular nitrogen compound metabolic process
1.82E-23
474
63
biosynthetic process
1.36E-13
388
42
neurotrophin TRK receptor signaling pathway
7.06E-13
44
44
protein binding transcription factor activity
1.83E-12
75
29
Fc-epsilon receptor signaling pathway
5.76E-12
32
24
protein complex
6.82E-11
385
64
gene expression
4.82E-10
70
35
cellular protein modification process
7.10E-10
232
41
molecular_function
7.10E-10
1552
66
extracellular matrix disassembly
1.72E-09
26
14
viral process
1.82E-09
60
49
symbiosis, encompassing mutualism through parasitism
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion.
The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition.
To identify the biological relevance of COVID-19 targeting human cellular miRNAs, KEGG pathway annotation, and GO analysis was performed on target gene pools. KEGG annotation data showed that COVID-19 targeting human miRNAs plays an important role in various cancer signaling, Hippo signaling pathways, cardiac (e.g cardiomyopathy, Adrenergic signaling in cardiomyocytes), cell cycle, FoxO and number of other signaling pathways (Table.5). Furthermore, GO pathway annotation analysis showed that the COVID-19 targeting human miRNAs involved in important signaling dysregulated during age-related complications, such as biosynthesis, metabolic, cellular protein modification, and cellular component assembly. Most importantly, we also found that signaling related to immune response, Fc-epsilon receptor signaling pathway, viral process, and symbiosis, encompassing mutualism through parasitism signaling affected by the COVID-19 targeting human cellular miRNAs (Table.6). The signaling mentioned above is important to fight against viral infections [45-48]. Both KEGG and GO pathway analysis revealed that COVID-19 targeting human cellular miRNAs are involved in the number of age-related complications. Impaired GO and KEGG pathways may be due to low abundance miRNA and might be one of the contributing factors for the increasing severity and mortality of the COVID-19 infection in aged individuals and with other underlying conditions.
The dual role of host cellular miRNAs cannot be denied in the viral replication or inhibition. Host miRNAs can have both functions, antiviral, which is beneficial to host, or miRNA-viral genome interaction can increase stability and beneficiary for viral propagation [18, 20, 22]. In a viral infection, we hypothesized that host cellular miRNAs predominantly have antiviral activity. Though possible, positive regulation of viral genome translation by miRNAs is less likely. Our data and hypothesis are based on in silico analysis and realize that not all miRNAs targets will be real. Considering the many miRNAs targets identified in our in-silico analysis, even if 5% of total miRNAs targets are real (based on target score), it is possible that these could be used for therapeutic purposes. Further, in vitro and in vivo studies will be needed to validate some of these targets. However, in view of the current lack of effective treatment strategies, innovative approaches need to be considered.
Supplementary Materials
Competing interests
The author declares no competing interests.
Acknowledgments
We would like to thank The Department of Medicine for their support. This publication is based upon work supported in part by the National Institutes of Health AG036675 (National Institute on Aging-AG036675 S.F, and C.I). The above-mentioned funding did not lead to any conflict of interest regarding the publication of this manuscript.
Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72?314 Cases From the Chinese Center for Disease Control and Prevention
CDC COVID-19 Response Team, Severe Outcomes Among Patients with Coronavirus Disease2019 (COVID-19) — United States, February 12-March 16, 2020, in Morbidity and Mortality Weekly Report. 2020, March27, US Department of Health and Human Services/Centers for Disease Control and Prevention
Increasing expression of microRNA 181 inhibits porcine reproductive and respiratory syndrome virus replication and has implications for controlling virus infection
GaoL, HeRQ, WuHY, ZhangTT, LiangHW, YeZH, et al. (2018).
Expression Signature and Role of miR-30d-5p in Non-Small Cell Lung Cancer: a Comprehensive Study Based on in Silico Analysis of Public Databases and in Vitro Experiments
HatseS, BrouwersB, DalmassoB, LaenenA, KenisC, SchöffskiP, et al. (2014).
Circulating MicroRNAs as Easy-to-Measure Aging Biomarkers in Older Breast Cancer Patients: Correlation with Chronological Age but Not with Fitness/Frailty Status
Coronaviruses post-SARS: update on replication and pathogenesis
1
2009
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
Pneumonia outbreak associated with a new coronavirus of probable bat origin
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
Transmission of 2019-nCoV Infection from an Asymptomatic Contact in Germany. N Engl J Med
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N Engl J Med
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
China Medical Treatment Expert Group for Covid-19. Clinical Characteristics of Coronavirus Disease 2019 in China
1
2020
... Novel Coronavirus was identified near the end of 2019, presenting with a spectrum of symptoms including febrile illness, cough, severe pneumonia, and in some patients, death. The pathogen is now widely called Coronavirus disease 2019 (COVID-19) and has rapidly turned into a global pandemic since originally being identified in Wuhan, China. Coronaviridae is the family of single-stranded (+ssRNA), pleomorphic, enveloped RNA viruses that consists of the Coronavirus genus [1]. COVID-19 is also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) due to the genomic and symptomatic similarities with the SARS-CoV that caused an epidemic in 2002-2004 [2]. SARS-CoV-2 is predominantly spread by person-to-person contact via virally loaded respiratory droplets [3]. However, the transmission may also occur by touching contaminated surfaces and subsequently touching one's eyes, nose, or mouth [4, 5]. The incubation period, the period before the first symptoms present itself, is on average of 4-5 days, with 95% presenting within 12.5 days [6,7]. ...
Infection prevention and control during health care when novel coronavirus (ncov) infection is suspected
3
2020
... The World Health Organization (WHO) reported 2,920,905 confirmed infected, and 203,269 COVID-19 related deaths globally as of April 25th, 2020 [8]. Drastic measures to combat SARS-CoV-2 are warranted due to its contagiousness, viability, and higher death toll compared to its predecessor SARS-CoV. Although SARS-CoV had a significantly higher case-fatality (around 10%) [9], the virus was not viable enough to remain in the human population and never spread in the United States and other countries like SARS-CoV-2 [10]. SARS-CoV-2, on the other hand, appears to rapidly spread, which is contributing to its global spread and a significantly higher number of cases. ...
... Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Features, Evaluation and Treatment Coronavirus (COVID-19)
2
2020
... The World Health Organization (WHO) reported 2,920,905 confirmed infected, and 203,269 COVID-19 related deaths globally as of April 25th, 2020 [8]. Drastic measures to combat SARS-CoV-2 are warranted due to its contagiousness, viability, and higher death toll compared to its predecessor SARS-CoV. Although SARS-CoV had a significantly higher case-fatality (around 10%) [9], the virus was not viable enough to remain in the human population and never spread in the United States and other countries like SARS-CoV-2 [10]. SARS-CoV-2, on the other hand, appears to rapidly spread, which is contributing to its global spread and a significantly higher number of cases. ...
... In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Severe acute respiratory syndrome (SARS) Frewuently Asked Questions 2004
1
... The World Health Organization (WHO) reported 2,920,905 confirmed infected, and 203,269 COVID-19 related deaths globally as of April 25th, 2020 [8]. Drastic measures to combat SARS-CoV-2 are warranted due to its contagiousness, viability, and higher death toll compared to its predecessor SARS-CoV. Although SARS-CoV had a significantly higher case-fatality (around 10%) [9], the virus was not viable enough to remain in the human population and never spread in the United States and other countries like SARS-CoV-2 [10]. SARS-CoV-2, on the other hand, appears to rapidly spread, which is contributing to its global spread and a significantly higher number of cases. ...
Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72?314 Cases From the Chinese Center for Disease Control and Prevention
4
2020
... Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... ]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... ]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
4
2020
... Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
... ]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
... ]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
MicroRNA-141-3p Negatively Modulates SDF-1 Expression in Age-Dependent Pathophysiology of Human and Murine Bone Marrow Stromal Cells
1
2019
... Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
Role of MicroRNA-141 in the Aging Musculoskeletal System: A Current Overview
0
2019
microRNA expression patterns reveal differential expression of target genes with age
0
2010
Genome-wide miRNA signatures of human longevity
2
2012
... Severe and critical cases of COVID-19 disproportionately affect middle-aged and older/aged populations, with increased mortality in aged adults [11]. Illness severity ranges from asymptomatic, mild, severe, and critical. Mild illness is characterized as no to mild pneumonia [11]. The severe disease presents with dyspnea, hypoxia, or >50% lung involvement, and critical illness occurs when there is respiratory failure, shock, or multiorgan failure [11]. The Centers for Disease Control and Prevention (CDC) and the WHO estimate the mortality rates of SARS-CoV-2 to be around 3.4% and case fatality rates (CFR) may increase up to 10-27 % in individuals 80 years old or older [8, 12]. SARS-CoV-2 is a novel virus, and very little is known about it. In such a scenario, it is crucial to understand the COVID-19 pathobiology to identify the innovative treatment strategy. Our laboratory and others have reported that miRNAs play a critical role in age-related complications [13-16]. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Increasing expression of microRNA 181 inhibits porcine reproductive and respiratory syndrome virus replication and has implications for controlling virus infection
3
2013
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
... ]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
microRNA-122 stimulates translation of hepatitis C virus RNA
3
2008
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
... The dual role of host cellular miRNAs cannot be denied in the viral replication or inhibition. Host miRNAs can have both functions, antiviral, which is beneficial to host, or miRNA-viral genome interaction can increase stability and beneficiary for viral propagation [18, 20, 22]. In a viral infection, we hypothesized that host cellular miRNAs predominantly have antiviral activity. Though possible, positive regulation of viral genome translation by miRNAs is less likely. Our data and hypothesis are based on in silico analysis and realize that not all miRNAs targets will be real. Considering the many miRNAs targets identified in our in-silico analysis, even if 5% of total miRNAs targets are real (based on target score), it is possible that these could be used for therapeutic purposes. Further, in vitro and in vivo studies will be needed to validate some of these targets. However, in view of the current lack of effective treatment strategies, innovative approaches need to be considered. ...
Cellular microRNAs inhibit replication of the H1N1 influenza A virus in infected cells
3
2010
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... ]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... ]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
Masking the 5' terminal nucleotides of the hepatitis C virus genome by an unconventional microRNA-target RNA complex
2
2011
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... The dual role of host cellular miRNAs cannot be denied in the viral replication or inhibition. Host miRNAs can have both functions, antiviral, which is beneficial to host, or miRNA-viral genome interaction can increase stability and beneficiary for viral propagation [18, 20, 22]. In a viral infection, we hypothesized that host cellular miRNAs predominantly have antiviral activity. Though possible, positive regulation of viral genome translation by miRNAs is less likely. Our data and hypothesis are based on in silico analysis and realize that not all miRNAs targets will be real. Considering the many miRNAs targets identified in our in-silico analysis, even if 5% of total miRNAs targets are real (based on target score), it is possible that these could be used for therapeutic purposes. Further, in vitro and in vivo studies will be needed to validate some of these targets. However, in view of the current lack of effective treatment strategies, innovative approaches need to be considered. ...
MiR-342-5p suppresses coxsackievirus B3 biosynthesis by targeting the 2C-coding region
2
2012
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... , 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
Stabilization of hepatitis C virus RNA by an Ago2-miR-122 complex. Proc Natl Acad Sci U S A
2
2012
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... The dual role of host cellular miRNAs cannot be denied in the viral replication or inhibition. Host miRNAs can have both functions, antiviral, which is beneficial to host, or miRNA-viral genome interaction can increase stability and beneficiary for viral propagation [18, 20, 22]. In a viral infection, we hypothesized that host cellular miRNAs predominantly have antiviral activity. Though possible, positive regulation of viral genome translation by miRNAs is less likely. Our data and hypothesis are based on in silico analysis and realize that not all miRNAs targets will be real. Considering the many miRNAs targets identified in our in-silico analysis, even if 5% of total miRNAs targets are real (based on target score), it is possible that these could be used for therapeutic purposes. Further, in vitro and in vivo studies will be needed to validate some of these targets. However, in view of the current lack of effective treatment strategies, innovative approaches need to be considered. ...
Human microRNA hsa-miR-296-5p suppresses enterovirus 71 replication by targeting the viral genome
4
2013
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... , 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... , 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
... Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
Human miR-3145 inhibits influenza A viruses replication by targeting and silencing viral PB1 gene
1
2015
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
The microRNA miR-485 targets host and influenza virus transcripts to regulate antiviral immunity and restrict viral replication
1
2015
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication
1
2008
... MiRNAs play a vital role in the pathogenesis of various diseases, including viral infections, disease progression, and inhibition [17-23]. MicroRNAs are small noncoding RNAs, bind to 3'UTR of mRNA, and inhibit translation or induce degradation of mRNAs [19, 21, 23]. Recent studies suggested that host cellular miRNAs can directly target both viral 3’UTR and coding region of the viral genome to induce antiviral effect [19, 21, 23]. For example, the number of groups previously reported that host miRNAs (miR-323, miR-491, miR-485, miR-654, and miR-3145) bind to influenza PB1 gene coding region, degrade RNA and inhibit viral translation and reduce the accumulation of viral particles [19, 24, 25]. Furthermore, the host cellular miRNA-29a inhibit Human immunodeficiency virus type 1 (HIV-1) nef protein expression and thus, inhibit viral replication [26]. On the contrary, some groups also suggested the positive effect of host miRNAs on viral replication. For example, miR-122 binding to 3’ and 5’ UTR of hepatotropic virus RNA and increase viral RNA stability leads to viral propagation [18, 20, 22]. Based on the above reports, we did in silico analysis of miRNAs targeting SARS and COVID-19 (recent isolates from different regions) to understand the pathophysiology and identify novel therapeutic targets. ...
miRDB: an online database for prediction of functional microRNA targets
2
2020
... As previously mentioned above, miRNAs are known to target 3'UTR and coding sequences and prevent mRNA translation or degrade RNA. Keeping this in consideration, we used the whole viral genome sequence for miRNA target analysis. We used miRDB (http://www.mirdb.org/) software to identify novel human miRNAs targeting the COVID-19 viral genome [27, 28]. Furthermore, we used this data to correlate with the existing literature. ...
... Gene Ontology (GO) and KEGG signaling pathway analyses were performed on human microRNAs targeting the COVID-19 genome using DIANA-miRPath v 3.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php) [29]. We used miRNAs with a target score above 90 because these miRNAs have high probability of being real targets [27, 28]. ...
Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data
2
2019
... As previously mentioned above, miRNAs are known to target 3'UTR and coding sequences and prevent mRNA translation or degrade RNA. Keeping this in consideration, we used the whole viral genome sequence for miRNA target analysis. We used miRDB (http://www.mirdb.org/) software to identify novel human miRNAs targeting the COVID-19 viral genome [27, 28]. Furthermore, we used this data to correlate with the existing literature. ...
... Gene Ontology (GO) and KEGG signaling pathway analyses were performed on human microRNAs targeting the COVID-19 genome using DIANA-miRPath v 3.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php) [29]. We used miRNAs with a target score above 90 because these miRNAs have high probability of being real targets [27, 28]. ...
DIANA-miRPath v3.0: deciphering microRNA function with experimental support
1
2015
... Gene Ontology (GO) and KEGG signaling pathway analyses were performed on human microRNAs targeting the COVID-19 genome using DIANA-miRPath v 3.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php) [29]. We used miRNAs with a target score above 90 because these miRNAs have high probability of being real targets [27, 28]. ...
Cellular miR-130b inhibits replication of porcine reproductive and respiratory syndrome virus in vitro and in vivo
1
2015
... Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
Endogenous cellular MicroRNAs mediate antiviral defense against in?uenza A Virus
1
2018
... Host cellular miRNAs are known to play an antiviral role in the number of published studies [18-23]. In this study, we performed in silico analysis of human cellular miRNAs targeting SARS and COVID-19 (isolates) genome and identified some novel miRNAs. We identified number (558) of common human cellular miRNAs targeting both SARS and COVID-19 genome. Top 10 common miRNAs have a target score of ≥95 (target score between 99-95), and each miRNA contains more than at least ten sites in the targeting viral genome (Table 3), indicating possible antiviral property for coronavirus infection (for both SARS and COVID-19). It will be interesting to verify these miRNA in-vitro and in-vivo animal models to use as a therapeutic target in the future. Cocktail of multiple miRNAs mimics through the intranasal route will be useful in coronavirus infection. These human miRNAs targeting coronavirus can be useful to combat any future outbreaks. In a previous report, host cellular miRNAs-181 binds to the ORF-4 region at the viral genome of porcine reproductive and respiratory syndrome virus (PRRSV) to inhibit its replication [17]. One step further, Guo et al. (2013) delivered intranasal inhalation of miR-181 mimics to slow down the progression of PRRSV in an experimental porcine model [17]. Another study used intranasal inoculation of miR-130 mimic to protect piglets from lethal challenge of PRRSV [30]. Similarly, the intranasal administration of chemically modified five miRNA mimics protected mice from H1N1 viral infection [31]. ...
The establishment of reference sequence for SARS-CoV-2 and variation analysis
1
2020
... In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Complete Genome Sequence of a 2019 Novel Coronavirus (SARS-CoV-2) Strain Isolated in Nepal
0
2020
Structural, glycosylation and antigenic variation between 2019 novel coronavirus (2019-nCoV) and SARS coronavirus (SARS-CoV)
1
2020
... In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Investigation of microRNA expression in human serum during the aging process
1
2015
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Age-associated microRNA expression in human peripheral blood is associated with all-cause mortality and age-related traits
2
2018
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
... ]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
miR-15a-5p suppresses inflammation and fibrosis of peritoneal mesothelial cells induced by peritoneal dialysis via targeting VEGFA
2
2019
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Targeting the circulating microRNA signature of obesity
2
2013
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Potential Involvement of MiR-30e-3p in Myocardial Injury Induced by Coronary Microembolization via Autophagy Activation
2
2017
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
miR-23c suppresses tumor growth of human hepatocellular carcinoma by attenuating ERBB2IP
2
2018
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Expression Signature and Role of miR-30d-5p in Non-Small Cell Lung Cancer: a Comprehensive Study Based on in Silico Analysis of Public Databases and in Vitro Experiments
2
2018
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
The potential value of miR-1 and miR-374b as biomarkers for colorectal cancer
1
2015
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
PIK3C2A is a gene-specific target of microRNA-518a-5p in imatinib mesylate-resistant gastrointestinal stromal tumor
2
2016
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
... The most important and striking feature of COVID-19 is the increased case fatality rate in aged individuals. The CDC reports 45% of cases requiring hospitalization are patients ≥65 years old [12]. Furthermore, 80% of COVID-19 related deaths occur in these patients (≥65 years old) [12]. Moreover, individuals with underlying conditions such as cardiovascular, chronic lung disease, diabetes, kidney, liver diseases, and cancer are at higher risk for severe illness and mortality. On the contrary, <1% mortality is observed in patients between 20-54 years of age or younger [12]. Based on miRNA target analysis and published literature, we hypothesized that the COVID-19 genome replicates and propagates viral particles easily in the aged individuals compared to younger due to the overall low abundance of miRNA expression with age. Recent human studies reported that the overall expression of miRNAs levels goes down with age [16, 35, 36]. For example, Huan et al. (2018) reported that 81% (103 miRNAs) microRNAs were negatively, and 19% (24 miRNAs) positively correlated with age in human peripheral blood [36]. We speculate that in younger individuals with COVID-19 infection, host cellular miRNAs bind to the complementary site of the viral RNA genome and prevent its replication; however, in an aged individual, this mechanism is not that efficient leads to accumulation of viral particle and severe illness. This might be true for underlying conditions as well. Some of the miRNAs targeting the COVID-19 genome showed low abundance (down-regulated) with some underlying conditions (Table 7). For example, miR-15b-5p (Coronary artery disease) [37], miR-15a-5p (Kidney disease) [38], miR-520c-3p (obesity/diabetes) [39], miR-30e-3p (Myocardial Injury) [40], miR-23c (hepatocellular carcinoma) [41], miR-30d-5p (non-small cell lung cancer) [42], miR-4684-3p (colorectal cancer) 43], and miR-518a-5p (Gastrointestinal stromal tumors) [44] down-regulated in pathophysiological condition. ...
Coronavirus infections and immune responses
1
2020
... To identify the biological relevance of COVID-19 targeting human cellular miRNAs, KEGG pathway annotation, and GO analysis was performed on target gene pools. KEGG annotation data showed that COVID-19 targeting human miRNAs plays an important role in various cancer signaling, Hippo signaling pathways, cardiac (e.g cardiomyopathy, Adrenergic signaling in cardiomyocytes), cell cycle, FoxO and number of other signaling pathways (Table.5). Furthermore, GO pathway annotation analysis showed that the COVID-19 targeting human miRNAs involved in important signaling dysregulated during age-related complications, such as biosynthesis, metabolic, cellular protein modification, and cellular component assembly. Most importantly, we also found that signaling related to immune response, Fc-epsilon receptor signaling pathway, viral process, and symbiosis, encompassing mutualism through parasitism signaling affected by the COVID-19 targeting human cellular miRNAs (Table.6). The signaling mentioned above is important to fight against viral infections [45-48]. Both KEGG and GO pathway analysis revealed that COVID-19 targeting human cellular miRNAs are involved in the number of age-related complications. Impaired GO and KEGG pathways may be due to low abundance miRNA and might be one of the contributing factors for the increasing severity and mortality of the COVID-19 infection in aged individuals and with other underlying conditions. ...
Sensor Sensibility-HIV-1 and the Innate Immune Response
0
2020
Representing virus-host interactions and other multi-organism processes in the Gene Ontology
0
2015
Gene expression profiles alteration after infection of virus, bacteria, and parasite in the Olive flounder (Paralichthys olivaceus)
1
2018
... To identify the biological relevance of COVID-19 targeting human cellular miRNAs, KEGG pathway annotation, and GO analysis was performed on target gene pools. KEGG annotation data showed that COVID-19 targeting human miRNAs plays an important role in various cancer signaling, Hippo signaling pathways, cardiac (e.g cardiomyopathy, Adrenergic signaling in cardiomyocytes), cell cycle, FoxO and number of other signaling pathways (Table.5). Furthermore, GO pathway annotation analysis showed that the COVID-19 targeting human miRNAs involved in important signaling dysregulated during age-related complications, such as biosynthesis, metabolic, cellular protein modification, and cellular component assembly. Most importantly, we also found that signaling related to immune response, Fc-epsilon receptor signaling pathway, viral process, and symbiosis, encompassing mutualism through parasitism signaling affected by the COVID-19 targeting human cellular miRNAs (Table.6). The signaling mentioned above is important to fight against viral infections [45-48]. Both KEGG and GO pathway analysis revealed that COVID-19 targeting human cellular miRNAs are involved in the number of age-related complications. Impaired GO and KEGG pathways may be due to low abundance miRNA and might be one of the contributing factors for the increasing severity and mortality of the COVID-19 infection in aged individuals and with other underlying conditions. ...
Downregulation of exosome-encapsulated miR-548c-5p is associated with poor prognosis in colorectal cancer
1
2019
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
miR-548d-3p inhibits osteosarcoma by downregulating KRAS
1
2019
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
MiR-409-3p Inhibits Cell Proliferation and Invasion of Osteosarcoma by Targeting Zinc-Finger E-Box-Binding Homeobox-1
1
2019
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Circulating MicroRNAs as Easy-to-Measure Aging Biomarkers in Older Breast Cancer Patients: Correlation with Chronological Age but Not with Fitness/Frailty Status
1
2014
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
Downregulation of miR-505-3p predicts poor bone metastasis-free survival in prostate cancer
1
2019
... List of selected human miRNAs targeting the COVID-19 genome down-regulated with age and underlying conditions.
miRNA
Decrease Expression in age related diseases (Human)
Reference
miR-15b-5p
Coronary Artery Disease
Zhu et al 2017 [37]
miR-15a-5p
Kidney disease
Shang et al 2019 [38]
miR-548c-5p
Colorectal Cancer
Peng et al 2018 [49]
miR-548d-3p
Osteosarcoma
Chen et al 2019 [50]
miR-409-3p
Osteosarcoma
Wu et al 2019 [51]
miR-30b-5p
Plasma (Aging)
Hatse et al 2014 [52]
miR-505-3p
Prostate cancer
Tang et al 2019 [53]
miR-520c-3p
Obesity/diabetes
Ortega et al 2013 [39]
miR-30e-3p
Myocardial Injury
Wang et al 2017 [40]
miR-23c
Hepatocellular carcinoma
Zhang et al 2018 [41]
miR-30d-5p
Non-small cell lung cancer
Gao et al, 2018 [42]
miR-4684-3p
Colorectal cancer
Wu et al, 2015 [43]
miR-518a-5p
Gastrointestinal tumors
Shi et al, 2016 [44]
In humans, coronavirus infection is pervasive and usually causes common cold-like symptoms, but some of them can lead to serious illness. Previous coronavirus outbreak in 2002, known as SARS-CoV spread worldwide but was contained quickly [9]. On the other hand, the current coronavirus (COVID-19) is highly contagious and spread worldwide rapidly and became a global pandemic [8,11]. It will be interesting to know what separates these two viruses based on the genomic sequence. Ours (Table 2 and Fig. 1) and other data [32-34] of SARS-CoV and COVID-19 genome sequence analysis showed sequence homology of around ~78.7%, which was obvious, but our miRNA target scan data showed striking results. We found considerable changes in the number of human miRNAs targeting SARS-CoV (848 miRNAs) and COVID-19 (~873 miRNAs). Moreover, we also found unique miRNAs for SARS-CoV (290 miRNAs) and COVID-19 (315 miRNAs) isolates. At nucleotide (Table 2) and phylogenetic levels (Fig. 1), both viruses look closely related, but host cellular miRNAs target comparison differences are much more significant. This might be one of the reasons for COVID-19 to increase infectivity and easy viral propagation compared to SARS-CoV. The COVID-19 recent isolates (twenty-nine different isolates) are closely related at nucleotide, phylogenetic (Fig. 1 and Table 2), and host cellular miRNAs target level (Fig. 2 and Supplementary Table 2). The Netherland (EPI_ISL_422601), Australia (EPI_ISL_413214), and Wuhan (EPI_ISL_403931) isolates have more mutation and few different host cellular miRNAs targeting sites compared to other 26 COVID-19 isolates (Table 4). In our study, we used 29 isolates from different geographical reagion, which informs that COVID-19 nucleotide sequences and thus the human miRNAs target sites are not changing among the isolates/viruses (Table 4). In most of the COVID-19 cases, the virus entered a country from different geographical locations. At the time of writing this paper, only the USA and China had sequenced multiple viruses isolates from a different region. There is a need to sequence more of the COVID-19 viral genome from different parts of each country before coming to any conclusion. ...
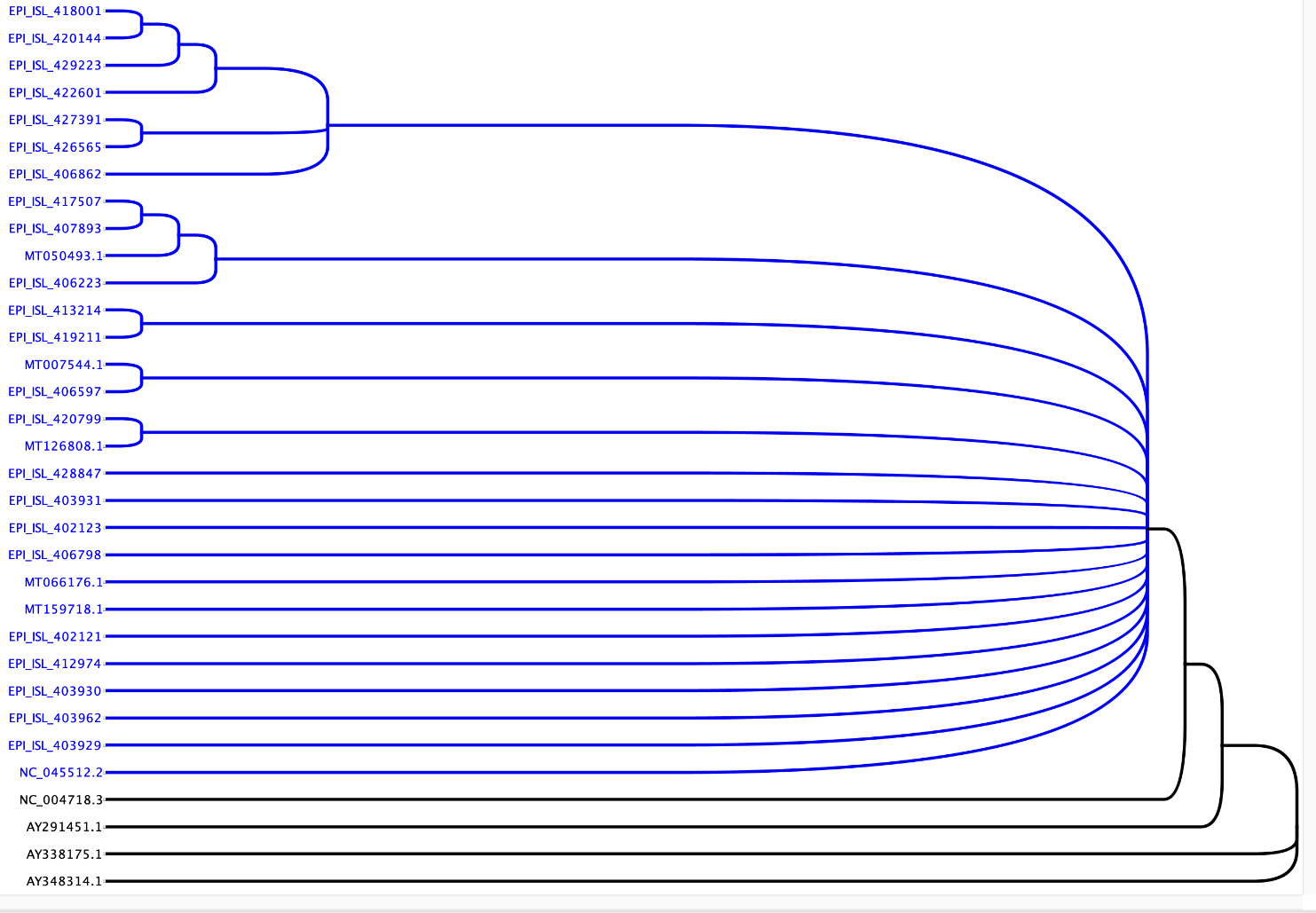

{kind=link}
{kind=link}
{kind=link}
{kind=link}



